2.1 理解分子遗传学和遗传力对研究人类行为至关重要
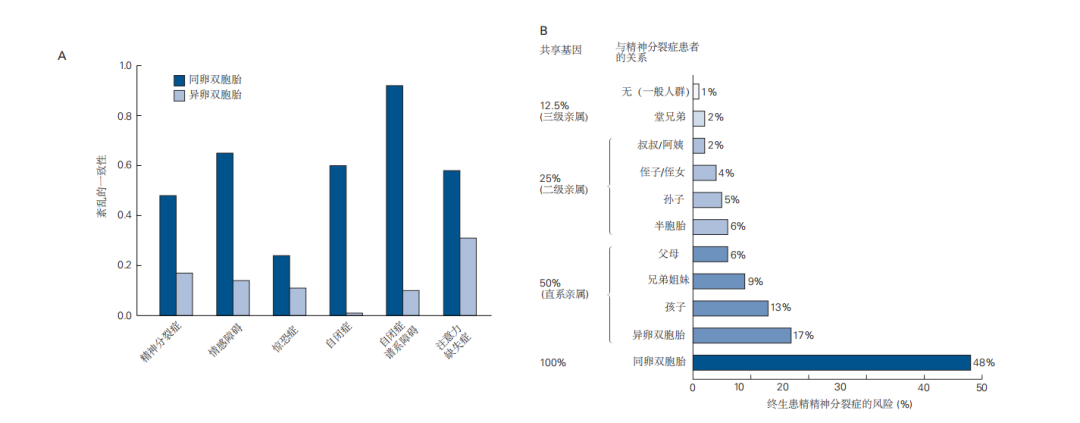
图2.1.1 精神疾病的家族风险提供了遗传性的证据。A. 同卵双胞胎之间的精神疾病相关性显著高于异卵双胞胎。同卵双胞胎几乎共享所有基因,因此他们患相同疾病的风险也较高(虽然不是100%)。异卵双胞胎则共享约50%的遗传物质。图中的“0”表示完全没有相关性(相当于两个随机个体之间的平均结果),而“1.0”表示完全相关。B. 精神分裂症患者的近亲罹患精神分裂症的风险更高。与异卵双胞胎类似,父母、子女和兄弟姐妹共享50%的遗传物质。如果精神分裂症由单一基因引发,那么患者的父母、兄弟姐妹、子女和异卵双胞胎的患病风险应相同。但家庭成员之间的风险差异表明,更复杂的遗传和环境因素共同作用于疾病的发生。
2.2 对基因组结构和功能的理解正在不断发展
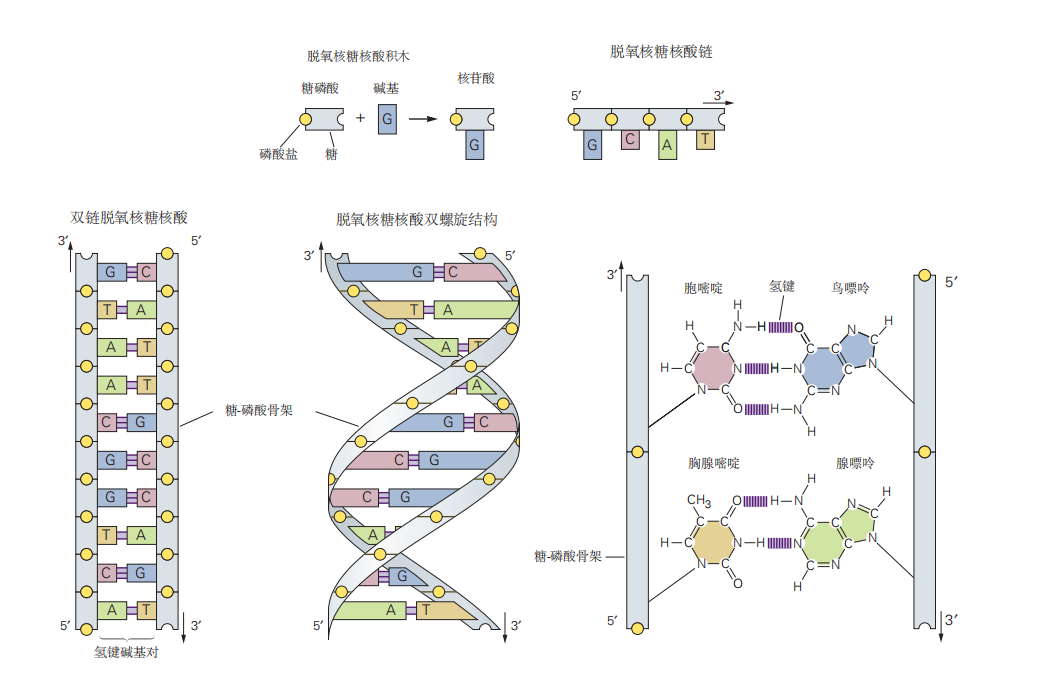
图 2.2.1:DNA的结构。DNA由双螺旋结构组成,四种核苷酸碱基——腺嘌呤、胸腺嘧啶、胞嘧啶和鸟嘌呤——沿着糖磷酸骨架排列在双链的DNA螺旋上。
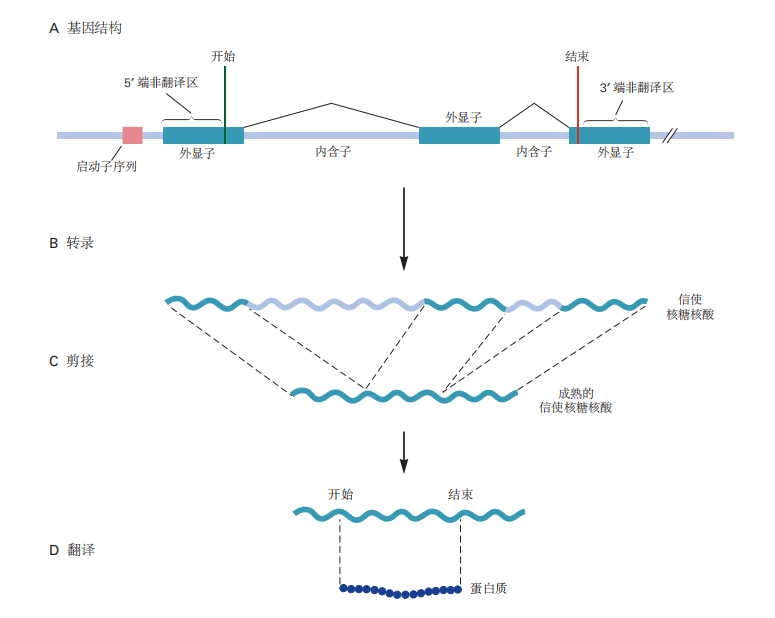
图 2.2.2:基因结构与表达。A. 基因由编码区(外显子)和非编码区(内含子)组成,内含子将外显子分隔开。基因的转录受非编码区的调控,例如启动子和增强子,这些通常位于基因的起始位置附近。B. 转录过程生成包含外显子和内含子的初级单链RNA转录本。C. 剪接过程将内含子从未成熟的转录本中移除,并将外显子连接形成成熟的mRNA,后者随后从细胞核中输出。D. 成熟的mRNA在翻译过程中生成蛋白质产物。
2.2.1 基因在染色体上的排列
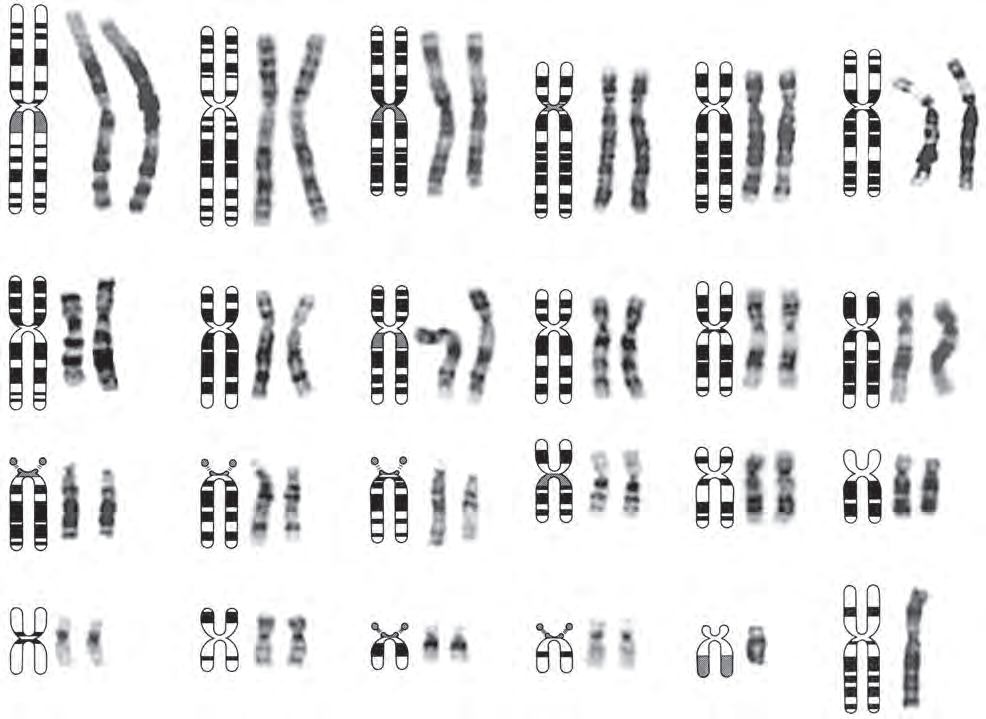
图 2.2.3:中期阶段的人类染色体图解,展示了每条染色体的独特形态。染色体的特定大小以及其上的明暗条带图案,使得每条染色体可以彼此区分。
2.3 基因型与表型之间的关系通常非常复杂
尽管DNA复制通常具有很高的准确性,但自发的错误——称为突变——仍然会发生。突变可能由于嘌呤和嘧啶碱基的损伤、DNA复制过程中的错误或减数分裂中的重组所引发。
在编码区内,单个DNA碱基的变化(也称为点突变)可分为五大类:
1.无声突变(silent mutation):改变了碱基,但不会导致蛋白质的功能发生明显变化。
2.错义突变(missense mutation):导致蛋白质中的一个氨基酸被另一个取代。随着信息学技术和实验手段的发展,这些突变可以进一步细分为两类:损害蛋白质功能的突变和对功能无明显影响的突变。
3.无义突变(nonsense mutation):将编码特定氨基酸的密码子(三联核苷酸)转变为终止密码子,导致蛋白质产物缩短(截短)。
4.剪接位点突变(canonical splice site mutation):改变了外显子/内含子边界的核苷酸,影响RNA剪接。
5.框移突变(frameshift mutation):由于插入或缺失小片段核苷酸,导致阅读框的改变,从而产生截短或异常的蛋白质。现有文献中,后四类突变(包括破坏性错义突变)通常被称为可能破坏基因的突变。
在实验遗传学研究中,化学诱变剂或电离辐射的暴露会显著增加突变频率。化学诱变剂倾向于诱导点突变,涉及单个DNA碱基对的变化或几个碱基对的缺失。电离辐射则可能导致大范围的插入、缺失或易位。
在人类中,点突变以较低的自发率发生于卵子和精子中,导致子代出现突变,而父母则没有,这种突变被称为新生突变。每代人中,整个基因组(约30亿碱基对)会发生70至90个单碱基变化,其中一个碱基对平均会导致一个编码蛋白质的基因发生错义突变或无义突变。父亲年龄越大,子代新生突变的数量越多,而母亲年龄较大时,子代出现染色体异常的概率更高。
自2001年人类基因组测序以来,随着检测基因变异的高分辨率方法不断提升,现在我们知道,点突变并不是人类之间唯一的序列差异。某些序列可能在染色体上缺失或多次重复,因此不同个体可能具有不同数量的基因拷贝。超过一个碱基但少于1000个碱基对的变化被称为插入缺失突变。当这种变异超过1000个碱基对时,它们被称为拷贝数变异。
任何基因变异对疾病或综合征的贡献可分为简单(或孟德尔)遗传和复杂遗传。通常,简单或孟德尔突变足以引起特定的表型,而不需要额外的遗传风险因素。这并不意味着所有携带突变的人都会表现出完全相同的表型,但通常特定疾病等位基因与表型之间的关系高度一致,接近一对一的关系(例如镰状细胞性贫血或亨廷顿舞蹈症)。
相反,复杂遗传疾病涉及多个遗传风险因素,它们改变了疾病发生的概率,但并非完全决定疾病发生。这种遗传贡献可能包括罕见突变、常见多态性或两者兼有,并且通常具有高度异质性,多个不同的基因和等位基因可能增加风险或提供保护。大多数复杂疾病也与环境因素密切相关。
2.4 基因在进化中的保守性
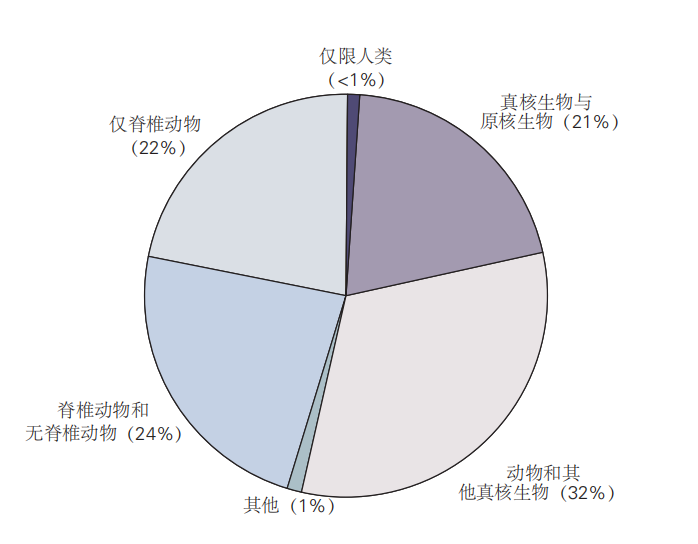
图 2.4.1: 大多数人类基因与其他物种的基因存在关联。只有不到1%的人类基因是人类所特有的;其余基因则可能在所有生物、所有真核生物、动物或仅脊椎动物中共享。
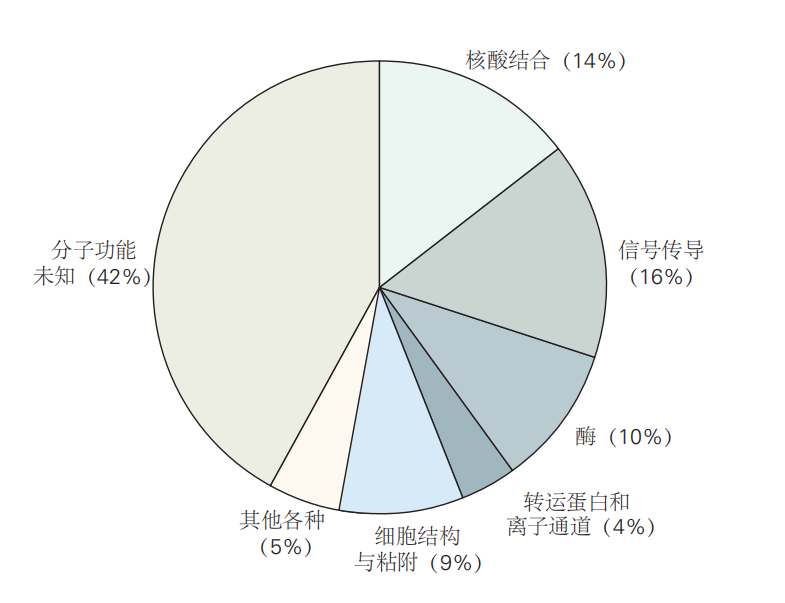
图 2.4.2: 26,383个人类基因的预测分子功能分类。
2.5 在动物模型中研究行为的遗传调控
2.5.1 转录振荡器调节苍蝇、小鼠和人类的昼夜节律
果蝇的随机诱变
果蝇行为的遗传分析通常在经过基因突变的果蝇个体中进行。突变可以通过化学诱变或插入诱变来实现,这些方法能够影响基因组中的任何基因。类似的随机诱变策略也被应用于秀丽隐杆线虫、斑马鱼和小鼠中,以产生突变体。
化学诱变,例如使用甲磺酸乙酯,通常会在基因中引发随机点突变。当可移动的DNA序列(称为转座子)随机插入到其他基因时,就会发生插入突变。
在果蝇中,最广泛使用的转座元件是P元件。P元件可以被修饰为携带眼睛颜色的遗传标记,这使得它们在遗传杂交中易于追踪。同时,P元件还可以被修饰以改变它们所插入基因的表达。
为了激活P元件的转位,携带P元件的果蝇菌株与不携带P元件的菌株杂交。遗传杂交会导致后代中P元件的不稳定和移位,最终P元件在随机基因中转移到新的位置。
小鼠的靶向诱变
随着哺乳动物基因操作技术的进步,科学家可以用突变版本精确替换已知的正常基因。产生突变小鼠菌株的过程涉及两个步骤。首先,利用同源重组在胚胎干细胞中特定的染色体基因上进行替换;然后,将修饰后的细胞系整合到胚胎的生殖细胞群中(见图2.5.1f)。
首先需要克隆目标基因并使其发生突变,然后引入选择性标记(通常是耐药性基因)到突变片段中。将改变后的基因引入胚胎干细胞,并分离出包含这些改变基因的细胞克隆。对每个克隆的DNA样本进行测试,以确定突变基因已整合到特定的同源位点而不是其他随机位点。
一旦鉴定出合适的细胞克隆,便在胚泡阶段(受精后3至4天)将这些细胞注射到小鼠胚胎中。胚胎被重新植入经过激素准备的雌性体内,允许其正常发育。得到的胚胎是由干细胞系和宿主胚胎混合形成的嵌合体。
小鼠的胚胎干细胞有能力参与发育的各个方面,包括生殖系。注射的细胞可以成为生殖细胞,并将改变后的基因传递给后代小鼠。这项技术已被广泛用于研究与神经系统发育或功能相关的各种基因。
限制基因敲除与调控转基因表达
为了提高基因敲除技术的应用效果,研究人员开发了将基因缺失限制在特定组织或发育阶段的技术。区域限制的一种方法是使用Cre/loxP系统。Cre/loxP系统是一种来源于P1噬菌体的位点特异性重组系统,其中噬菌体酶Cre重组酶催化34bp loxP识别序列之间的重组。这些序列可以通过同源重组插入到胚胎干细胞的基因组中,位于目标基因(称为floxed基因)的外显子两侧。当干细胞被注射到胚胎中时,最终可以培育出含有floxed基因的小鼠,这些基因在所有细胞中仍然活跃。
接下来,可以产生表达Cre重组酶的第二批转基因小鼠,它们在神经启动子序列的控制下表达Cre酶,这些序列通常只在大脑的特定区域中表达。通过将这两种小鼠杂交,感兴趣的基因只会在表达Cre转基因的细胞中被删除。
例如,在图2.5.2A中,编码N-甲基-D-天冬氨酸(NMDA)受体NR1亚基的基因被loxP元件夹住,然后将这种基因与在钙/钙调蛋白依赖性蛋白激酶2启动子控制下表达Cre重组酶的小鼠杂交。这个启动子通常在前脑神经元中表达,因此导致海马CA1区NR1亚基的选择性缺失(见图2.5.2B)。由于钙/钙调蛋白依赖性蛋白激酶2启动子只在出生后激活基因转录,这种策略可以最大限度地减少早期发育变化的影响。
除了区域限制外,对基因表达时间的控制为研究人员提供了更大的灵活性,并能够排除转基因引起发育缺陷的可能性。这可以通过构建可由药物控制开启或关闭的基因来实现。
这种方法的第一步是创建两个小鼠品系。品系1携带一个特殊的转基因,该转基因受四环素反应元件启动子的控制,而该启动子通常只存在于细菌中。这个启动子本身无法启动基因表达;它需要由特定的转录调节因子激活。因此,第二组小鼠表达另一种转基因,该转基因编码一种杂交转录因子——四环素激活因子(tTA),它能识别并结合四环素反应元件启动子。tTA的表达可以置于小鼠基因组中的特定启动子控制下,该启动子通常只在特定类别的神经元或大脑区域中激活基因转录。
当这两种小鼠交配时,一些后代将同时携带这两种转基因。在这些小鼠中,tTA与四环素反应元件启动子结合并激活下游的转基因表达。tTA特别有用,因为它在与四环素类抗生素结合时会失活,从而通过给小鼠服用抗生素来调节转基因的表达。还可以创建表达一种称为反向四环素激活因子(rtTA)的突变形式的小鼠。在这种情况下,转基因通常是关闭的,除非给小鼠喂食抗生素多西环素(见图2.5.3)。
核糖核酸干扰和CRISPR改变基因功能
最后,研究人员还可以使用现代分子工具靶向并灭活基因。一种方法是核糖核酸干扰(RNAi),该方法利用了真核细胞中大多数双链RNA被常规降解的机制。即使RNA只有一部分是双链的,整个RNA也会被降解。通过引入一个短的RNA序列,人为地使选定的mRNA形成双链,研究人员可以激活这一过程,从而降低特定基因的mRNA水平。
通过将DNA注射到新受精卵的细胞核中,可以在小鼠体内实验性地引入基因(见图2.5.4)。在一些注射的卵子中,新的基因或转基因会随机整合到染色体上的某个位置。由于胚胎处于单细胞阶段,这个整合的基因会在细胞分裂过程中被复制,最终进入动物的所有(或几乎所有)细胞,包括生殖细胞。
例如,通过将用于产生色素的基因注射到来自白化病菌株的受精卵中,可以恢复小鼠的外观颜色。有色素斑块的小鼠表明DNA成功整合并表达。通过测试注射后动物的DNA样本,可以确认转基因的存在。
在果蝇中也使用了类似的方法。研究人员将待注射的DNA克隆到可转座元件(P元件)中。当注射到胚胎中时,这些DNA被插入到生殖细胞核的DNA中。P元件可以被设计为在特定时间和特定细胞中表达基因。转基因可以是恢复突变体功能的野生型基因,或者是设计用来改变其他基因表达或编码特定变异蛋白质的基因。
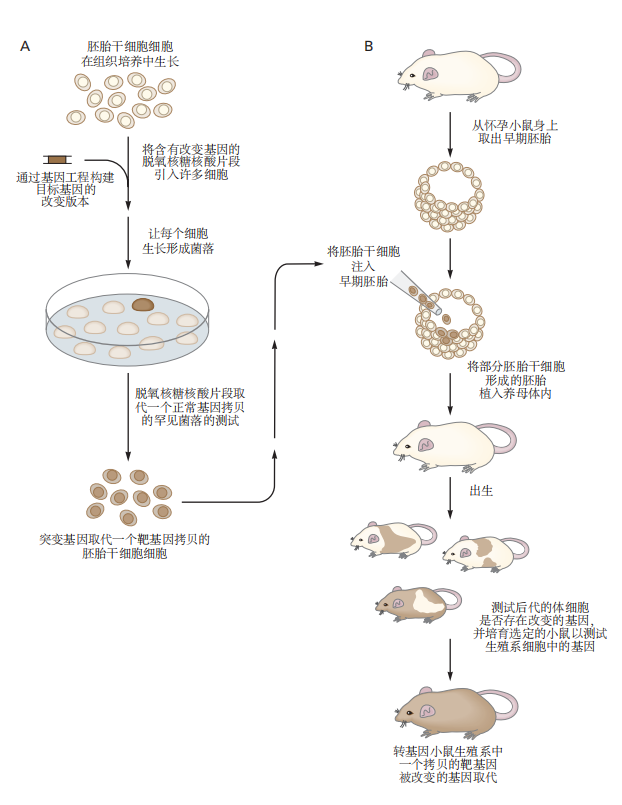
图 2.5.1: 突变小鼠菌株的生成过程。
A. 生成具有特定靶向突变的小鼠胚胎干细胞。
B. 利用经过基因修改的胚胎干细胞来创造转基因小鼠。
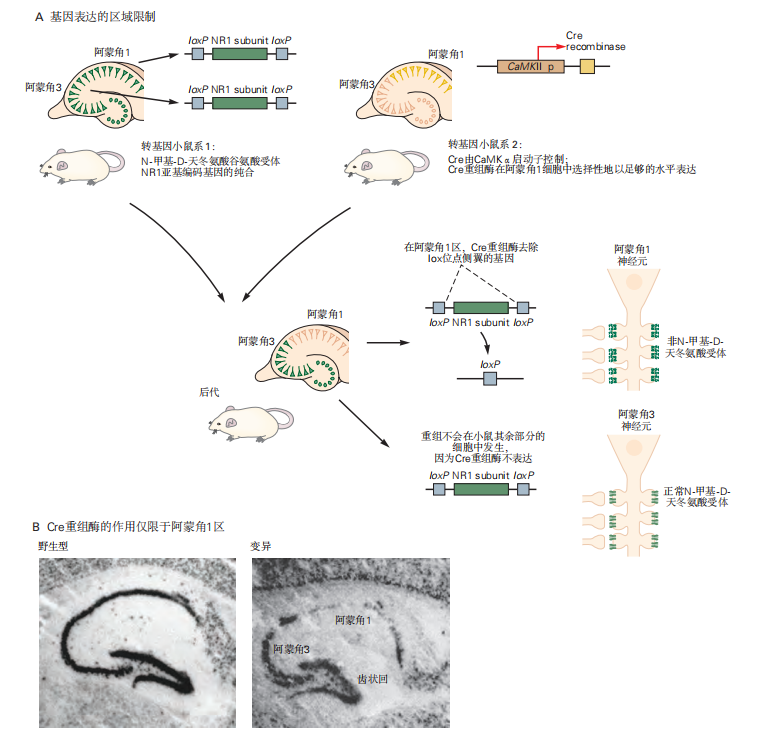
图 2.5.2: Cre/loxP 系统用于选择性区域基因敲除。
A. 培育了一种小鼠品系,其中编码 N-甲基-D-天冬氨酸受体 NR1 亚基的基因被 loxP 元件包围(转基因小鼠品系 1)。这些“floxed NR1”小鼠随后与第二种小鼠品系杂交,该品系在特定细胞或组织类型中表达 Cre 重组酶(转基因小鼠品系 2)。在这个例子中,Cre 基因的表达由钙/钙调蛋白依赖性蛋白激酶 2a 启动子驱动。在携带 Cre 重组酶转基因且为 Floxy 基因纯合的后代中,Cre 介导的 loxP 重组将删除 Floxy 基因,且这一过程仅发生在表达 Cre 的细胞类型中。
B. 通过原位杂交检测野生型和突变小鼠海马切片中 NR1 亚基的 mRNA 表达情况。结果显示,突变小鼠的海马阿蒙角 1 区中 NR1 的 mRNA 表达(暗染色)显著减少,而在阿蒙角 3 和齿状回区则保持正常。
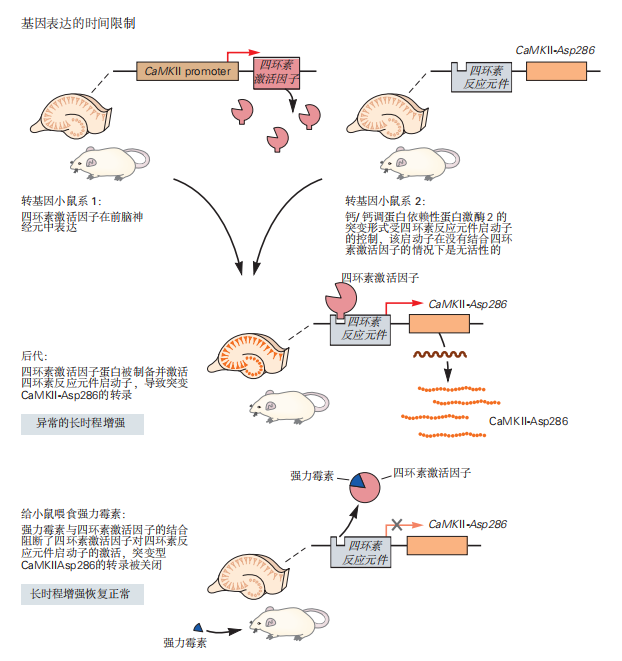
图 2.5.3: 四环素系统用于转基因表达的时间和空间调控。
培育了两个独立的转基因小鼠品系。一个品系在钙/钙调蛋白依赖性蛋白激酶 2a 启动子的控制下表达四环素激活因子,这是一种经过工程改造的蛋白质,能够结合并识别细菌四环素反应元件。第二个品系携带一个受四环素反应元件控制的感兴趣的转基因(在此例中为一种组成型活性的钙/钙调蛋白依赖性蛋白激酶 2),这种激酶在没有 Ca2+ 的情况下仍能保持活性。当这两个品系交配后,后代的小鼠在前脑中以特定模式表达四环素激活因子蛋白。当四环素激活因子蛋白与四环素反应元件结合时,便会激活下游转基因的表达。通过给这些小鼠服用四环素(或多西环素),可以使四环素激活因子蛋白发生构象变化,解除与四环素反应元件的结合,从而阻断转基因的表达。通过这种方式,小鼠可以在前脑中表达钙/钙调蛋白依赖性蛋白激酶 2–Asp286,并通过施用多西环素来控制这种表达。
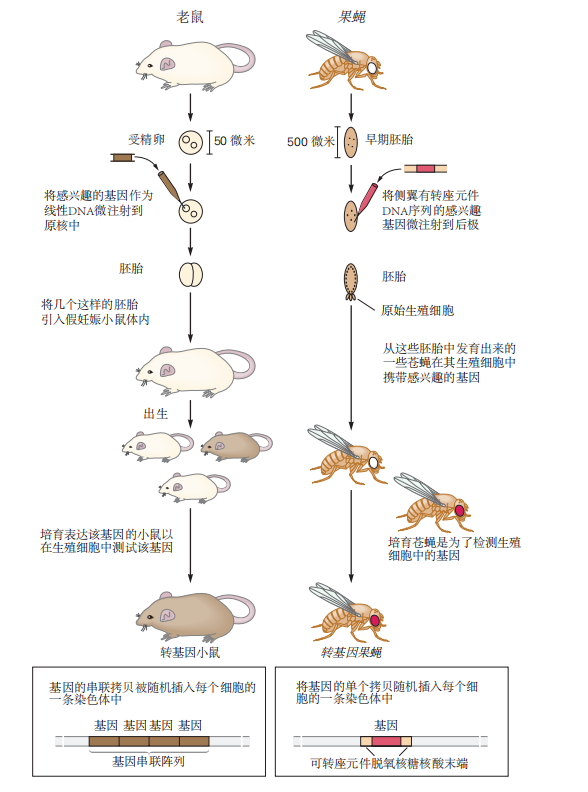
图 2.5.4: 转基因小鼠和果蝇的生成。
在此,注射到小鼠体内的基因会引起毛色的变化,而注射到果蝇体内的基因则会引起眼睛颜色的变化。在这两种物种的一些转基因动物中,DNA 被插入到不同细胞的不同染色体位点(见底部插图)
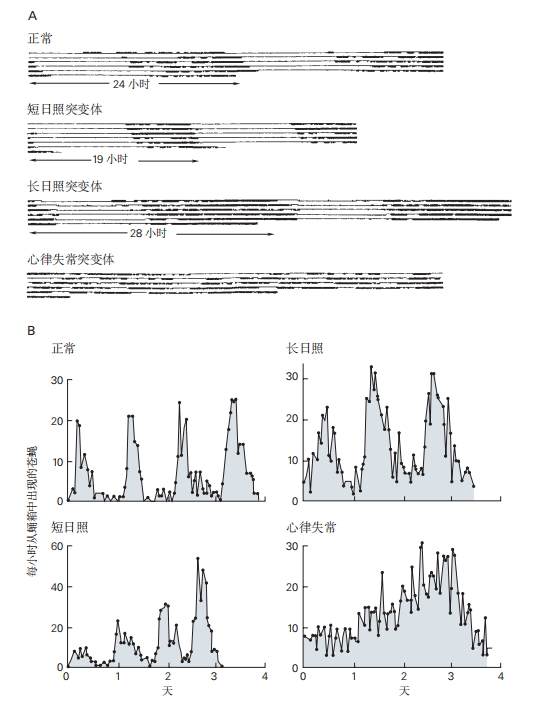
图 2.5.5:单个基因控制果蝇行为的昼夜节律。per 基因的突变会影响果蝇内部生物钟调节的所有昼夜节律行为。
A. 正常果蝇和per 基因突变体(短日照、长日照和无节律)三种品系的活动节律。在将果蝇从12小时光照和12小时黑暗的循环环境转移到持续黑暗后,通过红外光监测它们的活动。记录中的粗线段表示活动。
B. 正常成年果蝇群体以周期性的方式从蛹壳中出现,即使在持续黑暗中也是如此。图中显示了在持续黑暗的4天期间,每小时出现的果蝇数量(来自四个不同群体)。无节律突变体群体在黑暗中没有表现出可辨别的节律。
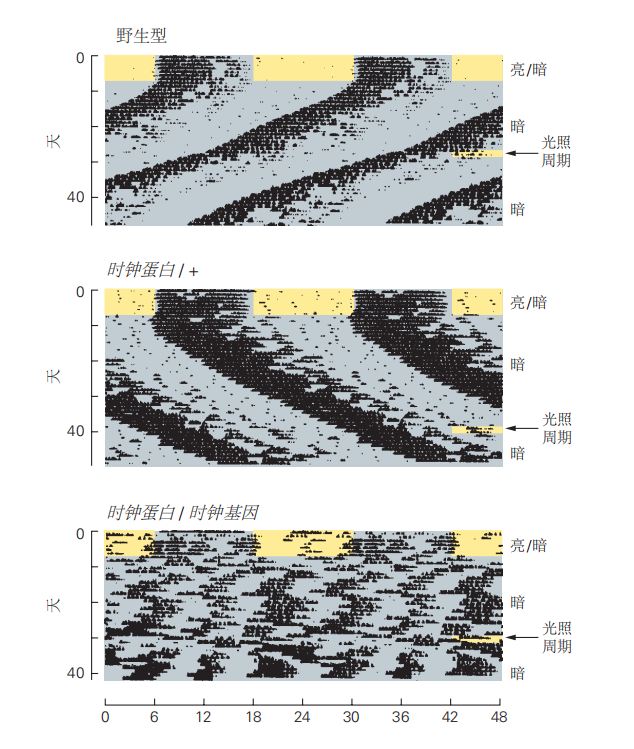
图 2.5.6:时钟基因对小鼠昼夜节律的调控。记录显示了三种类型小鼠的活动周期:野生型、杂合型和纯合型。所有小鼠在前7天经历12小时的光暗循环,然后转移到持续黑暗的环境中。之后,它们被暴露在6小时的光照周期中,以重置其节律。野生型小鼠的昼夜节律周期为23.1小时。杂合型时钟基因小鼠的周期为24.9小时,而纯合型时钟基因小鼠在转移到持续黑暗时完全丧失了昼夜节律,并在光照期后短暂表现出28.4小时的节律。
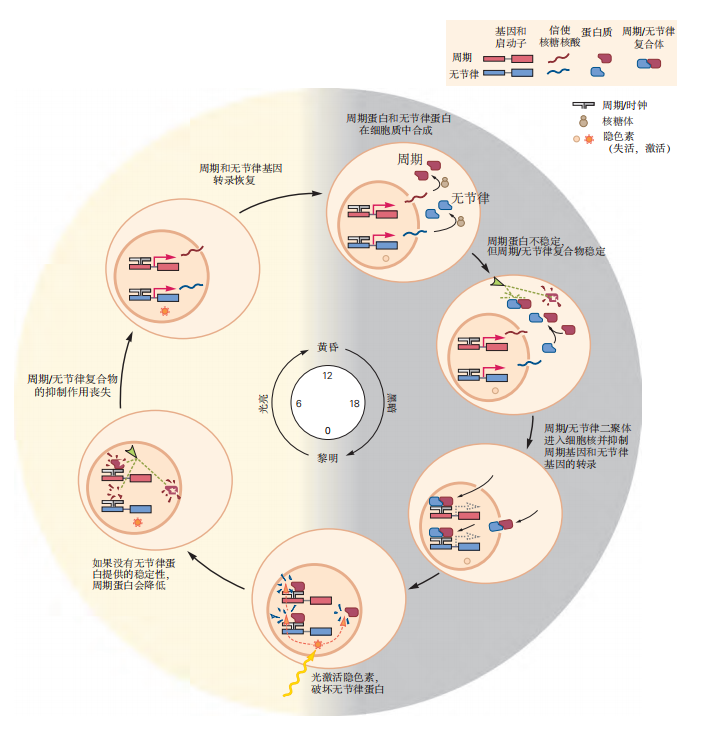
图 2.5.7:驱动昼夜节律的分子机制。生物钟基因受两种核蛋白——周期蛋白(PER)和无节律蛋白(TIM)的调节。这些蛋白质逐渐积累,并相互结合形成二聚体。一旦形成二聚体,它们便进入细胞核并关闭包括它们自身在内的昼夜节律基因的表达。这一过程通过抑制时钟蛋白(CLOCK)和CYCLE蛋白的作用来实现,它们是驱动节律基因和无节律基因转录的关键因子。周期蛋白非常不稳定,大部分在降解前未能抑制由时钟蛋白介导的基因转录。周期蛋白的降解由多种蛋白激酶通过磷酸化事件调控。当周期蛋白与无节律蛋白结合时,它们相互保护,避免降解。随着时钟蛋白驱动更多节律基因和无节律基因的表达,足够的周期蛋白和无节律蛋白最终积累并形成稳定的二聚体,进入细胞核抑制它们自身的转录。结果,节律基因和无节律基因的mRNA水平下降;随后,周期蛋白和无节律蛋白的水平也下降,时钟蛋白可以再次驱动节律基因和无节律基因的表达。在白天,无节律蛋白通过光调节信号通路(包括隐花色素)降解,因此周期蛋白/无节律蛋白复合物只在夜间形成。时钟蛋白诱导周期蛋白和无节律蛋白的表达,但这些蛋白质随后会抑制时钟蛋白的功能。
2.5.2 蛋白激酶的自然变异调节果蝇和蜜蜂的活动
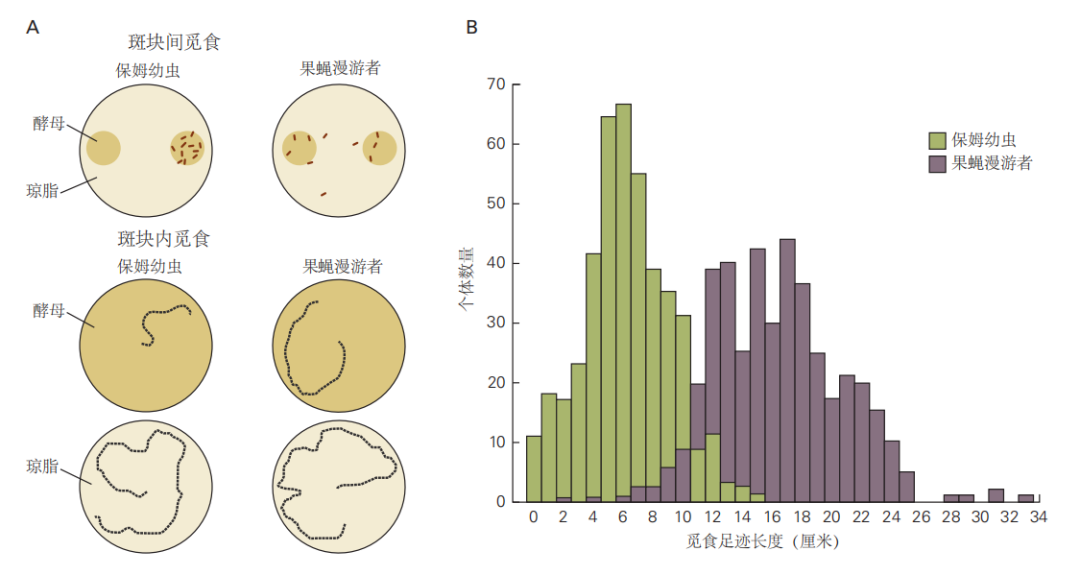
图 2.5.8:索科洛夫斯基通过不同野生果蝇的杂交实验,研究了“漫游者”和“守家者”幼虫之间的遗传差异。结果表明,这种差异主要由一个基因决定,即觅食基因座。觅食基因编码一种信号转导酶,这种酶由细胞代谢物环鸟苷-3,5-单磷酸(cGMP)激活。因此,这种自然行为差异源于信号转导途径的调控变化。许多神经元功能受蛋白激酶调节,例如由觅食基因编码的cGMP依赖性激酶。像蛋白激酶这样的分子在将短期神经信号转化为神经元或神经回路的长期特性变化中起着关键作用。
2.5.3 神经肽受体调节几种物种的社会行为
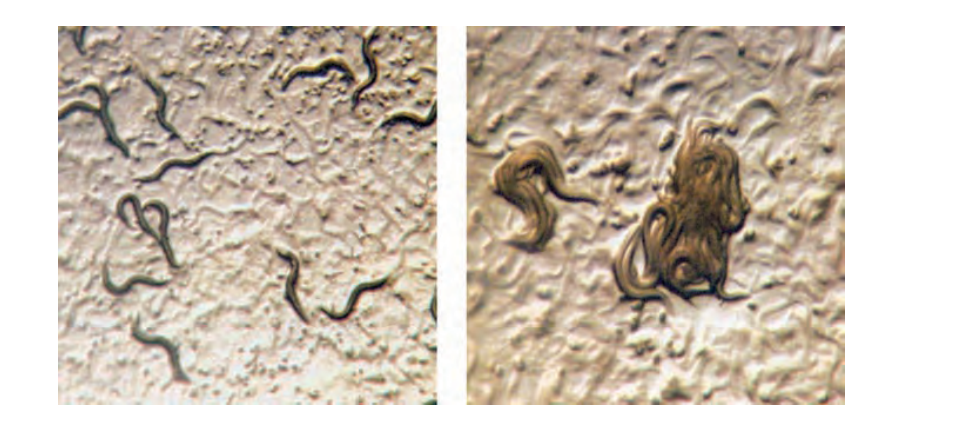
图 2.5.9:秀丽隐杆线虫的进食行为取决于编码神经肽受体的基因活性。在一个品系中,个体蠕虫在孤立状态下单独觅食(左图),而在另一个品系中,个体则倾向于聚集在一起觅食。这种差异可以通过神经肽受体基因中的单个氨基酸替换来解释。
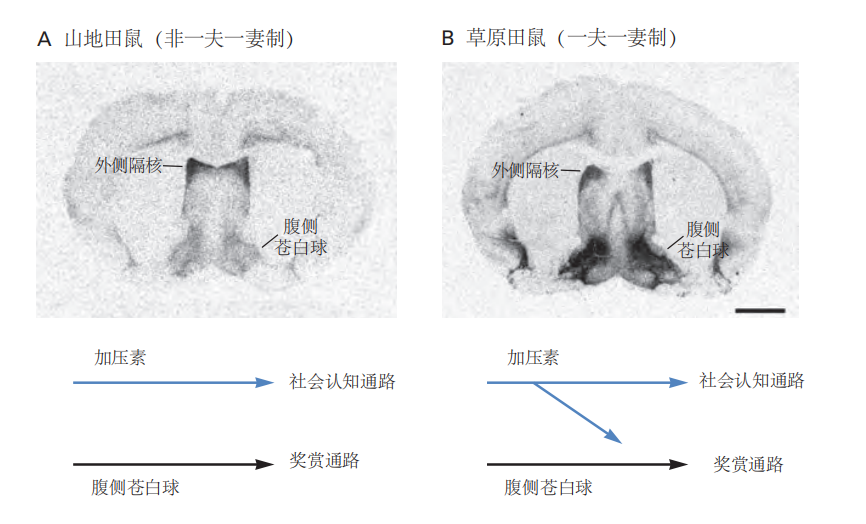
图 2.5.10:血管加压素受体在两种亲缘关系密切的啮齿动物中的分布差异。
A. 在山地田鼠的外侧隔核中,受体表达较高,但在腹侧苍白球中表达较低,这种模式不形成配对键。
B. 在一夫一妻制的草原田鼠中,受体在腹侧苍白球中的表达较高。受体在腹侧苍白球中的表达使血管加压素能够将社会认知通路与奖赏通路连接起来。
本文翻译整理自Principles of Neural Science, Sixth Edition,翻译初稿来自于GitHub/OpenHUTB,本公众号进行进一步校对排版后发布,仅用于学习交流使用,如需购买原版书籍可点击阅读原文购买。
— THE END —
来源 | Principles of Neural Science, Sixth Edition
图文排版 | 李芋岭
审核修改 | 王坤
(转载请保留此部分内容)

关注本号~

加入读者交流群~
(添加请备注单位姓名)

加入知识星球~
点亮赞与在看
让更多人看到
本篇文章来源于微信公众号:OncoLab