25.1 引言
癌症免疫监视,即免疫系统搜索并识别体内癌症和癌细胞的过程,其历史可以追溯到20世纪初,当时保罗·埃尔利希就预测免疫系统可以抑制肿瘤的生长。
此后,免疫学家对癌症免疫监视的概念进行了数十年的阐述和争论。在经历了许多希望和挫折之后,现在的实验证据明确地表明,这种免疫反应确实在宿主抵御癌症起至关重要的作用。
与此相关的有趣现象是,先天性免疫系统和适应性免疫系统有助于传统化疗和放疗的抗肿瘤效果,而这些效果被认为至少部分是通过在治疗过程中暴露于肿瘤抗原而激活先天性免疫介导的。因此,这些经典的癌症疗法与宿主的抗肿瘤免疫反应密切相关。
开发新的有效方法,利用免疫系统的力量来治疗癌症,已成为一个极具吸引力的热门研究领域。近年来,肿瘤免疫治疗取得了显著进展,包括肿瘤靶向单抗、免疫细胞检查点治疗、肿瘤疫苗和嵌合抗原受体(CAR)T细胞治疗。
虽然这些疗法在某些情况下,为许多类型的恶性肿瘤带来了显著的、持久的应答率,但受益的患者数量仍然有限。
因此,能够增强宿主抗肿瘤免疫反应且不良反应最小或可控的新疗法和现有疗法的组合仍然是该领域的重点。
先天性免疫系统在宿主抵御病原体入侵的过程中采用了许多与消灭癌症或癌细胞相同的机制。或许在这些过程中研究最充分的细胞类型是自然杀伤(NK)细胞。
在抗癌先天免疫反应中,NK 细胞可直接杀死受病毒感染的细胞和癌细胞。事实上,NK细胞受损或缺乏NK细胞的动物模型显示出了各种类型癌症发病率的增加。
已知包括单核细胞和粒细胞衍生细胞在内的其他先天性细胞类型也能在肿瘤微环境(TME)内外积极或消极地调节抗肿瘤适应性免疫。在细胞免疫的同时,先天免疫受体通过信号转导在调节适应性免疫反应中发挥重要作用,因其在调节肿瘤发生中的作用增加了人们对这些受体的关注。
相对于 T 细胞受体和 B 细胞受体,先天性免疫受体是数量有限的种系编码受体,可识别病原体相关分子模式(PAMP)或称为损伤相关分子模式(DAMP)的内源性应激信号。因此,模式识别受体(PRRs)不像T细胞和B细胞那样具有特异性。
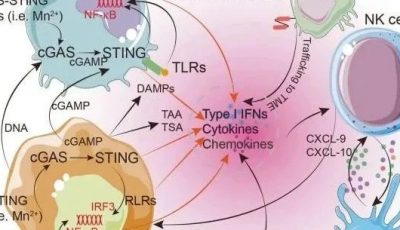
模式识别受体分为(1)膜相关PRRs,包括Toll样受体(TLRs)和C型凝集素受体(CLRs);(2)细胞质PRRs,包括RNA感受性视黄酸诱导基因-I(RIG-I)样受体(RLR)和DNA感受器;(3)可溶性PRRs,包括补体受体、胶原素、五角蛋白等。
前两类 PRRs(但不包括可溶性 PRRs)可以在与其同源配体结合时向细胞核传递信号,以改变细胞的基因表达谱。典型的情况是诱导 I 型干扰素(IFN)基因和炎症细胞因子基因,以介导抗微生物反应。
先天信号也可以直接驱动树突状细胞(DC)的成熟、抗原呈递以及细胞毒性CD8+T细胞的功能。
越来越多的证据表明,抗肿瘤免疫受 PRRs 激活的调控。
在动物模型中,外源性给予配体刺激PRRs可诱导强大的抗肿瘤免疫反应,而在某些PRRs缺陷小鼠模型中则观察到肿瘤过度生长。
另一方面,PRRs的激活引起炎症反应,于是提出了PRRs实际上是否可能阻碍癌症免疫的问题,因为某些类型的肿瘤的进展通常与炎症有关。
肿瘤细胞释放的内源性配体对先天PRRs的慢性激活可能通过促炎反应促进肿瘤进展,从而增强TME内的增殖、抗凋亡和促纤维化信号。
这些似乎相互矛盾的报道表明,先天免疫激活的PRRs具有双重作用,免疫增强作用,即增强抗肿瘤免疫反应,以及通过诱导慢性炎症促进肿瘤发生。
本章内容总结了我们对先天PRRs的了解,它们在肿瘤免疫监视和调控肿瘤发生中发挥作用。我们还讨论了利用先天性 PRRs 信号转导调节剂靶向肿瘤的潜在治疗意义。
25.1.1 TLRs 在调控肿瘤发生中的作用
到目前为止,TLR 家族受体是最容易理解的先天信号 PRRs 类群,它们是病原体感染的哨兵。TLRs可以识别微生物PAMPs,并通过募集同源接头蛋白来发出信号。
一般来说,TLRs 通常利用被称为髓系分化主要反应基因 88(MyD88)的适配体作为其信号传导途径;然而,TLR3 则利用含 TIR 结构域的适配体诱导 IFN-β(TRIF,又称 TICAM)信号传导途径,尽管有证据表明它也利用 MyD88 途径。TLR4同时需要MyD88和TRIF通路才能完全激活下游的信号级联。
在与TLRs结合后,接头蛋白与其他下游蛋白质结合,这些下游蛋白质介导转录因子和蛋白激酶的激活,如NF-kappaB(NF-κB)、干扰素调节因子(IRFs)和丝裂原活化蛋白激酶(MAPK),以调节靶基因的转录,如I型IFN和炎症细胞因子的转录。
TLRs在抗原提呈细胞中高度表达,一些TLRs的激活诱导抗肿瘤介质,如I型IFN,这促使人们努力利用 TLR 激动剂来治疗肿瘤。
事实上,TLRs是激活抗癌免疫反应的重要治疗靶点,最初因Coley毒素和卡介苗(BCG)相关的抗癌反应而被重视。
另一方面,TLR信号也诱导炎症反应,因此也可能有利于肿瘤的生长。
一个新兴的概念是,除了PAMP,TLRs还识别一系列被称为损伤相关分子(DAMPs)的自我衍生分子,它们在细胞应激时释放,可改变 TME。
因此,如下所述,TLRs明显地同时表现出了促肿瘤和抗肿瘤功能。这可能归因于不同的致瘤方案或小鼠类型,它们会影响肿瘤的发育过程和共生菌群。
TLR1/2/6这些TLR表达在细胞表面。
TLR2与TLR1或TLR6形成异二聚体,识别多种PAMP,包括脂肽、肽聚糖、脂磷壁酸、脂阿拉伯甘露聚糖、酵母多糖、克氏锥虫的糖基磷脂酰肌醇锚定的粘蛋白样糖蛋白以及细菌或酵母的血凝素蛋白。
TLR2还被证明能够识别内源性配体,如双聚糖、透明质酸、万西康和表面活性蛋白A。在动物模型中也已经进行了TLR2在肿瘤发生调控中的作用的研究,有证据表明TLR2信号可能发挥抗肿瘤或促肿瘤功能,其发挥的功能取决于所研究的模型。
在单次注射 N-亚硝基二乙胺(DEN)后重复注射四氯化碳(CCl 4)的肝细胞癌(HCC)模型(以下称 DEN/CCl 4 模型)中,TLR2 缺陷小鼠表现出肿瘤生长加剧。研究表明,包括干扰素-γ表达受损、细胞衰老和丧失自噬在内的抗肿瘤免疫反应的减弱是肿瘤生长扩大的原因。
同样,另一项研究表明,在DEN/CCL4模型中,TLR2缺陷小鼠会罹患更多更大的肠道肿瘤。肿瘤发展的特征是肠道TME中IL-6、IL-17A的增加以及信号转导和转录激活剂3(STAT3)磷酸化水平的增加。
另一方面,在STAT3过度激活的小鼠胃癌模型中,TLR2缺乏导致肿瘤负荷减少,表明TLR2具有促肿瘤作用。这种作用与炎症无关,其特征是宿主胃上皮细胞的增殖受损和凋亡增加。这些现象的解释是由于TLR2缺乏而抑制了这些细胞中的磷脂酰肌醇-3激酶(PI3K)、丝氨酸/苏氨酸激酶(AKT)、细胞外信号调节蛋白激酶(ERK1/2)、c-jun氨基末端激酶(JNK)和NF-κB通路。
此外,TLR2信号也可能在维持正常干细胞和胃癌细胞的“干性”方面发挥作用。在肺癌模型中,TLR2缺陷小鼠与野生型(WT)小鼠相比,LLC细胞的转移生长的速度较慢。WT小鼠肿瘤生长的增强可能是由于LLC衍生的Verscan激活TLR2:TLR6复合体,导致髓样细胞分泌可促进肿瘤生长的肿瘤坏死因子-α(TNF-α)。
TLR3 TLR3定位于内体中,识别来自病毒的内吞双链RNA(DsRNA),以及从死亡细胞中释放的衍生mRNA。
越来越多的证据表明,TLR3具有促进抗肿瘤免疫的作用。在前列腺癌植入性转基因的腺癌模型中,TLR3缺陷小鼠的肿瘤生长速度比对照组快。在这个模型中,TLR3-I型干扰素信号通路增强了NK细胞的抗肿瘤反应。
与此一致的是,一些研究已经检验了合成的TLR3配体聚肌苷胞酸(PolyI:C)在癌症治疗中的潜在疗效。PolyI:C-TLR3轴的可能作用机制是:(A)诱导DC细胞上依赖IRF3的NK激活分子(INAM),(B)肿瘤浸润性巨噬细胞向M1表型偏移,以及(C)激活DC,包括产生I型IFN以增强有效的细胞毒性T细胞免疫反应。
另一方面,TLR3也在自发的肺转移模型中促进肿瘤的生长,TLR3缺陷小鼠在该模型中的肺转移受到抑制。
进一步研究表明,来自肿瘤外切体的RNA可激活TLR3在肺上皮细胞中的表达,导致中性粒细胞募集和转移前生态位的发展,从而促进肿瘤的进展。
如上所述,几十年来TLR3激动剂Poly I:C一直被认为是一种有前途的癌症免疫治疗佐剂。这种疗法虽然有效,但已被证明会引起危及生命的副作用,如引发细胞因子风暴。在这方面,最近的一项研究报告了一种新型的合成RNA,旨在选择性地激活TRIF途径,从而有效地激活荷瘤小鼠的NK细胞和细胞毒性T细胞,而不会引发由其他类型的dsRNA诱导的严重细胞因子风暴。
TLR4 TLR4在细胞表面表达并识别革兰氏阴性杆菌的主要组分之一脂多糖(LPS)。
据报道,TLR4识别多种内源性配体,如高迁移率族蛋白1(HMGB1)、热休克蛋白(HSPs)、双糖链蛋白聚糖、透明质酸片段和氧化型低密度脂蛋白。
在单剂量偶氮甲烷(AOM)联合硫酸葡聚糖钠(DSS)联合作用的小鼠结肠癌模型中(以下称为AOM/DSS模型),TLR4缺陷小鼠显示出肿瘤负荷减少,表明TLR4具有促肿瘤作用。
从机制上讲,TLR4信号(可能由共生细菌激活)在结肠上皮细胞中诱导(A)免疫抑制环氧化酶-2(COX-2),(B)双调节素激活表皮生长因子受体(EGFR)信号转导,以及(C)通过募集表达COX-2的巨噬细胞形成TME。
根据上述结果,在肠上皮细胞中携带的绒毛蛋白启动子上具有构成活性的TLR4蛋白的转基因小鼠在同一小鼠模型中更容易发生肿瘤。然而,在另一个结肠癌模型中,TLR4显示出抗肿瘤作用。
在自发性肠道肿瘤发生的APCmin/+小鼠模型(APCmin模型)中,肠道上皮细胞中具有固有活性的TLR4的小鼠显示出肿瘤负荷减少。从这些小鼠的肠道分离的肿瘤细胞显示IFN-β的表达增加和半胱天冬蛋白酶-3的激活,这与体内细胞凋亡的增加有关。
在肝细胞癌模型中,即DEN/CCL4模型中,与WT小鼠相比,TLR4缺陷小鼠的肿瘤发展较慢,这暗示了TLR4信号的促肿瘤功能。
肠道的微生物区和肝脏细胞上的TLR4被认为间接的促进了增殖,阻止了肿瘤细胞的凋亡和肝脏有丝分裂原表皮生长素的表达。
同样,在DEN诱导的肝癌模型中,观察到TLR4缺陷小鼠的肝癌发展减少。然而另一项研究表明,在DEN诱导的肝癌模型中,TLR4缺陷小鼠表现出更高的肿瘤负荷。DNA的受损修复和随后的氧化应激可以解释这种癌变加剧的原因。
在7,12-二甲基苯并菲(DMBA)和巴豆油联合诱导的皮肤癌模型中,缺乏TLR4的小鼠也显示出抗癌能力,表明TLR4信号有促癌作用。从机理上讲,从濒死的角质形成细胞中释放出来的HMGB1,激活TLR4并增强炎症,从而促进肿瘤的发展。
此外,在黑色素瘤的基因工程小鼠模型中,即HGF-CDK4(R24C)小鼠,由于致癌 CDK4(R24C) 基因突变,肝细胞生长因子(HGF)受体酪氨酸激酶信号传导失调,细胞周期控制受损,TLR4 的缺乏可抑制紫外线诱导的肺转移增强。在这个实验设置中,TLR4被紫外线损伤角质形成细胞释放的细胞外HMGB1激活,导致中性粒细胞募集,诱导血管生成和黑色素瘤细胞向内皮细胞迁移。
另一方面,另一项研究表明,在DMBA皮肤癌模型中,TLR4缺陷小鼠的肿瘤负担增加,并伴随着血清IL-17水平的升高和干扰素-γ水平的降低,这表明Th1介导的抗肿瘤反应受损。
关于TLR4在其他器官中的作用的报道相当有限。在Kras G12D胰腺癌模型中,造血细胞室中缺乏TLR4的小鼠在P48CRE中胰腺上皮内瘤病变减少。
相反,TLR4缺乏会在注射3-甲基胆蒽和丁基羟基甲苯(BHT)后导致肺部肿瘤加重。TLR4也被证明对DMBA诱导的乳腺癌模型和4T1接种的转移模型都有保护作用。
TLR4信号的促肿瘤或抗肿瘤结果可能取决于MyD88和TRIF信号通路的激活状态,这两个信号通路分别具有促肿瘤和抗肿瘤生长的作用。
TLR5 TLR5在细胞表面表达,且能识别细菌鞭毛中的鞭毛蛋白。一项研究报道,肿瘤细胞异位表达鞭毛蛋白可诱导可检测到的针对EL4小鼠淋巴瘤的抗肿瘤免疫反应,从而将鞭毛蛋白用于肿瘤疫苗。这种作用是通过TLR5信号和由核苷酸结合的寡聚化结构域(NOD)包含类受体(NLR)激活而引起的炎症体来介导的。
然而,最近的一份报告表明,TLR5可以促进肿瘤的生长。在TLR5缺陷小鼠的肉瘤模型中显示出肿瘤生长受阻。TLR5 缺陷与 IL-6 表达减少有关,IL-6 表达减少会导致髓源性抑制细胞(MDSCs)招募减少,MDSCs 会导致γδ-T 细胞释放 galectin-1,从而抑制抗肿瘤适应性免疫反应并加速肿瘤进展。
有趣的是,依赖于 TLR5 的肿瘤生长加速是通过与共生微生物群的相互作用介导的。
TLR7/8 TLR7可识别来自RNA病毒的内吞体内的单链RNA(SsRNA)。它在浆细胞样树突状细胞(PDC)上高度表达,对于大量释放针对这种细胞类型的RNA病毒的I型IFN来说至关重要。人类TLR8也识别病毒单链RNA。然而,TLR8缺陷的小鼠细胞在产生针对病毒单链RNA的细胞因子方面却没有缺陷。TLR7还可以识别与自身抗体结合的自我衍生的单链RNA。
TLR7/8的小分子激动剂已被提议用作抗肿瘤免疫疗法药物。已知咪喹莫特通过激活TLR7-MyD88信号发挥抗肿瘤作用。一些研究已经阐明了该化合物抗肿瘤作用的详细机制。
例如,咪喹莫特使DC产生I型IFN,并通过肿瘤坏死因子相关凋亡诱导配体(TRAIL)和颗粒酶B的活性,将自己转化为能够直接清除肿瘤细胞的细胞。
此外,具有特定序列的免疫刺激 RNA 寡核苷酸也能以 TLR7 依赖性方式通过 NK 细胞诱导抗肿瘤免疫反应。
然而,遗传学研究表明,TLR7 对肿瘤发生的作用恰恰相反。一项研究表明,在表达突变基因K-Ras的小鼠胰腺癌模型中,造血细胞中TLR7的缺乏会阻止肿瘤的发展。另一项研究表明,在LLC肺癌转移模型中,TLR7 缺陷小鼠的肿瘤负荷较轻,生存期延长。
使用合成配体的研究和遗传研究之间的结果似乎不一致,因此关于TLR7信号对肿瘤进展的影响,目前还没有明确的解释;这可能取决于合成配体和内源性配体之间TLR7激活的幅度和持续时间,其他PRRs也可能如此(见下文)。
TLR9 内体中的TLR9识别未甲基化的CpG DNA基序,这些基序经常在病毒和细菌基因组中被发现。TLR9 还能以类似于 TLR7 的方式识别与自身抗体结合的自身 DNA。
TLR9信号在抗肿瘤免疫中的作用已被大量报道所强调。
最值得注意的是,TLR9配体CpG-寡核苷酸(CpG-ODN)对肿瘤生长的治疗作用已被广泛研究。CpG-ODN在治疗C3小鼠宫颈癌模型和C26小鼠结肠癌模型中诱导了显著的抗肿瘤作用,这种治疗所导致的肿瘤消退和延长生存需要CD8+T细胞的参与。
CpG-ODN对小鼠神经母细胞瘤细胞株Neur2a也有抑制作用,这似乎是由NK细胞介导的。
此外,在小鼠异种移植模型中,将另一种刺激 TLR9 的 ODN 与曲妥珠单抗(一种抗人表皮生长因子受体 2(HER2)的单克隆抗体)结合使用,可有效控制对曲妥珠单抗耐药的人类乳腺癌细胞。
有趣的是,这种ODN在膜水平上调节了TLR9与HER受体的相互作用,从而抑制HER所依赖的生长信号。
另一方面,TLR9可能促进肿瘤生长。与WT小鼠相比,原位移植携带K-RAS和P53突变基因的胰腺癌细胞在TLR9缺陷小鼠中显示出生长延迟。这种效应是胰腺TME特有的,因为在TLR9缺陷小鼠中,皮下移植的胰腺细胞没有显示出生长减少。
在机制上,TLR9激活胰腺星状细胞(PSCs)导致趋化因子(C-C基序)配体11(CCL11)的产生,通过其受体趋化因子(C-C基序)受体3(CCR3)导致肿瘤细胞的增殖。
此外,研究还表明,PSCs可能是通过CCL3-CCR5轴将调节性T细胞(Tregs)以依赖TLR9的方式募集到肿瘤周围部位。此外,免疫细胞中的 TLR9 也会将 MDSC 招募到肿瘤组织、器官和组织中,从而进一步加剧肿瘤的进展。
25.1.2 细胞质核酸感应器在肿瘤发展中的作用
胞质核酸感应器PRRs几乎在所有类型的细胞中表达,并通过检测RNA和DNA或它们的模拟物来激发先天性免疫反应。这些胞质PRRs激活的标志是I型IFNs的诱导,这种诱导是 PRRs 识别病毒衍生核酸从而产生有效抗病毒反应的基础 。由于I型IFNs的抗肿瘤功能已被广泛认识,这些PRRs受体在抗肿瘤免疫中的作用也一直是人们关注的焦点。
RLRs RIG-I(也称为DDX58)和黑色素瘤分化相关基因5(MDA5)能感知 RNA 病毒的复制中间体 dsRNA,从而在感染细胞中产生大量 I 型 IFNs。
该家族的另一个成员–LGP2作为RIG-I和MDA-5的负反馈调节因子,但在某些情况下可能促进MDA5信号通路。尽管这两种激活型的PRRs在总体结构上相似,但由于病毒衍生 RNA 的结构特征不同,它们检测的病毒种类也不同。RIG-I 与含有 5′-三磷酸的 ssRNA(如病毒 RNA 和体外转录的 dsRNA)特异性结合。
研究还表明,RIG-I 优先与短 dsRNA 结合,而 MDA5 则优先识别长 dsRNA。这两种 RLR 具有共同的信号特征。识别到 dsRNA 后,它们会被适配体 MAVS(又称 IPS-1、CARDIF 或 VISA)招募到线粒体外膜,从而激活多个转录因子,包括 IRF3、IRF7 和 NF-κB。IRF3 和 IRF7 主要介导 I 型 IFNs 的诱导,而 NF-κB 则调节炎症细胞因子的基因诱导(更多详情,请参阅 “细胞因子 “一节)。
这些RLRs的激活强劲诱导了I型IFN,促使许多研究人员评估它们在抗肿瘤反应中的作用,因为I型IFN可调节抗病毒反应之外的包括对细胞生长、分化和抗肿瘤免疫的影响等许多重要的细胞功能。
在人类肝癌组织中RIG-I的低表达预示着更差的预后和对干扰素-α治疗的更高抵抗力。RIG-I的肿瘤抑制作用已在RIG-I缺陷小鼠的肝癌模型中得到验证。另有报道称,RIG-I激活可诱导黑色素瘤细胞分泌细胞外囊泡(EV),并在其表面表达NKp30配体,从而触发NK细胞介导的黑色素瘤细胞对其的清除。
由于 MDA5 触发了与 RIG-I 类似甚至相同的信号通路,这直接意味着 MDA5 也参与了肿瘤的发展。
由于LGP2增强了CD8+T细胞的存活和激活,因此LGP2也可能在抗肿瘤免疫反应中发挥直接调节作用。
事实上,RIG-I 的激活还能通过 BH3 家族诱导细胞内在凋亡,而这种凋亡与 I 型 IFN 信号传导无关,因此人们开始研究如何利用 RIG-I 介导的细胞凋亡来选择性地消灭恶性细胞 。
因此,人们致力于开发 RLR 信号通路的靶向药物,用于癌症治疗,其工作假设是,I 型 IFN、细胞凋亡或这两种通路的诱导可能起着至关重要的作用。
DNA感应器 在迄今报道的DNA感应PRRs中,环GMP-AMP(cGAMP)合成酶(cGAS)在抗病毒免疫中的作用是特征最明显的分子之一。释放到细胞质中的病毒 DNA 在 cGAS 催化下转化为 cGAMP,cGAMP 反过来又与 IFN 基因刺激因子(STING)结合,激活其下游信号通路,包括 I 型 IFNs 。
一些研究表明cGAS参与了宿主对肿瘤的免疫反应。表达cGAS的巨噬细胞对于肿瘤来源的β反应产生干扰素-DNA是必不可少的。与此一致的是,照射的肿瘤细胞刺激DC释放IFN-β,以cGAS依赖的方式促进抗原特异性CD8+T细胞的激活。
此外,抗癌药物顺铂和喜树碱也需要 cGAS 才能触发 IFN-β 的产生。
除了cGAS介导的途径外,其他DNA传感机制对于胞质核酸的反应也很重要。
DNA依赖的IRFs激活剂(DAI)与TBK1和IRF3结合并应答胞内DNA,诱导I型IFN,其抗肿瘤作用已被报道。另一种DNA传感的PRR–人的IFN-γ诱导蛋白16(IFI16)及其小鼠同源基因p206在胞浆DNA刺激下诱导IRF3激活和IFN-β的产生,其中STING到IFI16的募集可能是信号通路中的关键步骤。
另一种PRR-DDX41也观察到了类似与STING之间的关联。DDX41识别多种DNA,并作为cGAMP的第二信使直接感应cGAMP-di-GMP和cGAMP。已有报道称在由 DDX41 基因突变引起的造血肿瘤中,DDX41 具有抑制肿瘤的作用 。
此外,一些DNA损伤反应分子,如DNA依赖的蛋白激酶(DNA-PK)和减数分裂重组11(Mre11),参与了胞质DNA传感;然而尚不清楚这些DNA传感器是否调控肿瘤的发展。
总体而言,由于肿瘤来源的DNA被TME中的抗原提呈细胞摄取并激活DNA传感的PRRs,其抗肿瘤功能一直是基础和临床癌症免疫学关注的焦点。
STING(干扰素基因刺激因子,stimulator of interferon genes, STING)在抗肿瘤免疫反应中的作用一直是人们特别关注的焦点。
在接种了表达免疫原肽的肿瘤细胞的小鼠中,STING似乎有助于对这些细胞的抗肿瘤反应,其中肿瘤来源的DNA被TME中的抗原提呈细胞摄取,刺激STING并诱导IRF3介导的IFN-β的产生,以扩增抗原特异性CD8+T细胞。
类似地,放射治疗的肿瘤也会触发STING依赖性的IFN-β的产生和CD8+T细胞的激活。这表明了,这种IFN-β的上调是由死亡细胞产生的DNA通过激活IRF3诱导的。
此外,STING促进肿瘤组织中IL-18和IL22BP的表达,并抑制AOM/DSS结肠癌的发生。
另一方面,据报道,STING信号通路可以下调促炎因子IL-6的表达,而IL-6可激活促肿瘤转录因子STAT3。
一项使用胶质瘤模型的研究表明,STING通过增强CD8+T细胞的活性和减少免疫抑制细胞(如MDSCs和Tregs)在大脑中的浸润来减轻相关的癌症负担。
总的来说,靶向激活的STING在许多小鼠模型中对癌症的治疗是有益的。给予cGAMP可减少结肠癌26细胞的肿瘤生长,与DC成熟有关。
此外,cGAMP还通过I型IFN信号途径激活TME(肿瘤微环境)中的CD8+T细胞,从而抑制体内B16黑色素瘤细胞的生长。
由化合物ML RR-S2激活的STING可以在几种小鼠模型中发挥抗肿瘤活性。ML RR-S2在实验上也可用作GM-CSF表达的死亡肿瘤细胞抗癌疫苗的佐剂。这种治疗策略被称为 STINGVAX,已被证明对检查点疗法耐药的 B16 小鼠黑色素瘤有效。
与此相关,在这方面也值得注意的是,一些抗癌化学药物也是DNA加合物形成剂,触发细胞死亡并释放STING激动剂。已发现5,6-二甲基黄原酮-4-醋酸(DMXAA)是一种血管干扰剂,可直接与STING结合,现已被用作STING激动剂用于肿瘤治疗。DMXAA诱导抗原提呈细胞产生IFNβ,并通过增强获得性免疫对体内肿瘤细胞的生长显示出强大的治疗活性。
此外,DMXAA 还能靶向巨噬细胞,并将其从 M2 型极化为 M1 型。DMXAA治疗促进了对肿瘤的免疫记忆,通过DMXAA治疗而获得了排斥某种肿瘤效应的小鼠对继发性肿瘤同样具有抵抗力。
值得注意的是,这种药物的瘤内给药抑制了其他远处部位的肿瘤生长。因此,STING激活分子,如胞浆环二核苷酸和化学化合物,可能为建立有效的癌症免疫治疗铺平道路。
另一方面,有报道显示STING具有促肿瘤作用。在致癌物损伤的细胞中,DNA释放到胞浆中会刺激STING诱导炎性细胞因子的表达,并加剧DMBA诱导的皮肤癌。
也有报道称,免疫抑制宿主体内的STING可促进LLC肿瘤细胞的皮内生长和肺转移。这项研究还表明,针对gp100转导的LLC细胞的抗肿瘤反应是以STING依赖的方式诱导的,这意味着STING介导的抗肿瘤或促肿瘤反应依赖于肿瘤细胞的免疫原性。
本文由Oncolab微信公众号翻译整理自Oncoimmunology: A Practical Guide for Cancer Immunotherapy,仅用于学习交流使用,如需购买原版书籍可点击阅读原文购买。
— THE END —
来源 | Oncoimmunology: A Practical Guide
for Cancer Immunotherapy
作者 | Sho Hangai, Yoshitaka Kimura,Tadatsugu Taniguchi, and Hideyuki Yanai
初稿翻译 | 温鑫
编辑排版 | 吕思贤
审核修改 | 王坤 胡珊
(转载请保留此部分内容)

关注本号~

加入读者交流群~
(添加请备注单位姓名)

加入知识星球~
点亮赞与在看
让更多人看到
本篇文章来源于微信公众号:OncoLab