我们在前两章中已经学习了肿瘤微环境以及肿瘤免疫杀伤的重要执行者-CD8+ T细胞,这这一章中我们将学习激活抗肿瘤特异性免疫的另一个必要条件-肿瘤突变表位,它也是人体适应性免疫系统识别肿瘤细胞的重要前提。
4.1 引言
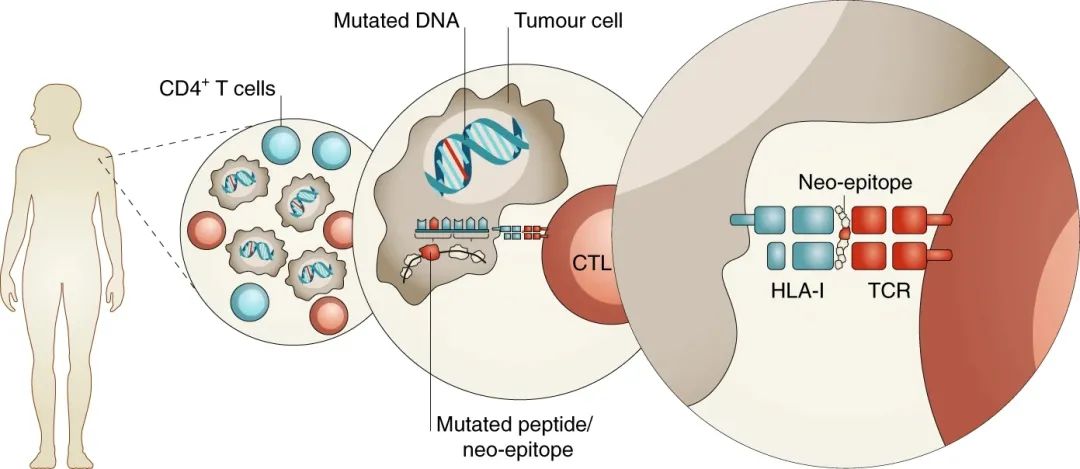
癌症的靶向免疫治疗是一个迅速发展的领域,其发展历史至少已经跨越了30年的历史。从肿瘤特异性抗原 (TSA)(与转化细胞完全相关的蛋白质分子)的鉴定和表征开始,以及最近由免疫检查点阻断疗法激活的新抗原特异性免疫细胞反应性的证据,证明了基于免疫的干预措施将在很大程度上塑造癌症治疗的未来。新抗原来自自然处理的宿主突变蛋白分子–最终以免疫原性肽的形式呈递给免疫系统。为了更好地了解诊断和治疗应用中的免疫学和生物学基础,增强癌症患者的医疗保健,对新抗原的产生和识别的更深入的了解是必不可少的。
在本章中我们简要地向读者介绍了癌症中的抗原的处理和呈递机制,并简要描述了癌症抗原的发现历史,以及一些开创性的发现。最后,我们讨论了癌症免疫治疗的最新进展,重点放在一些新抗原导向的治疗策略上,这些策略在临床转化和治疗的背景下可能会在未来的一段时间内得到改进。我们将本章的关注重点放在癌症患者的主动细胞治疗(ACT)上,以及将突变表位相关治疗策略与细胞治疗联合使用的可能性。
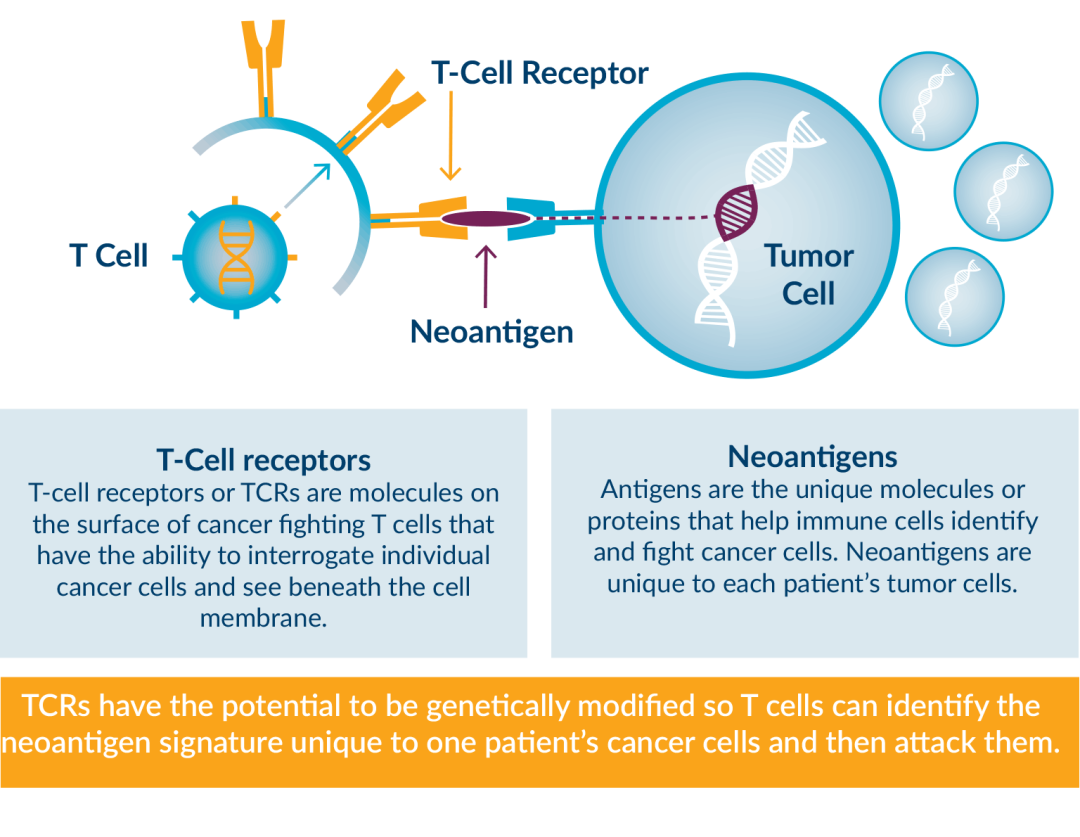
利用新抗原表位特异性 T 细胞亚群的潜力来杀伤肿瘤的方法非常有吸引力,因为它们能够识别和响应肿瘤细胞,具有较小的脱靶毒性以及非常高的效率,并且能够为癌症患者提供持久的和有临床意义的治疗结果。这种针对转化细胞的抗癌反应本质上是一种有针对性且富有成效的自身免疫反应,依赖于能够识别突变目标的T细胞受体(TCR)谱系的存在。
一些癌症抗原因其在恶性/转化细胞中的选择性组织表达或过度表达而被鉴定为“癌症抗原”,如间皮素或癌-睾丸抗原(将在本章后面讨论)。在其他情况下,发生在其他正常表达和功能蛋白中的突变可能会导致它们成为致癌因子。这些突变的宿主分子可能参与癌症的启动(致癌)、疾病持续或转移。由于某些突变对于恶性转化和肿瘤细胞存活至关重要,因此它们也可能促使免疫逃逸的发生,通过促进肿瘤的 T 细胞反应,或 通过诱导免疫功能丧失来“致盲”抗癌免疫反应,此二者并非相互排斥,反而相互适应。
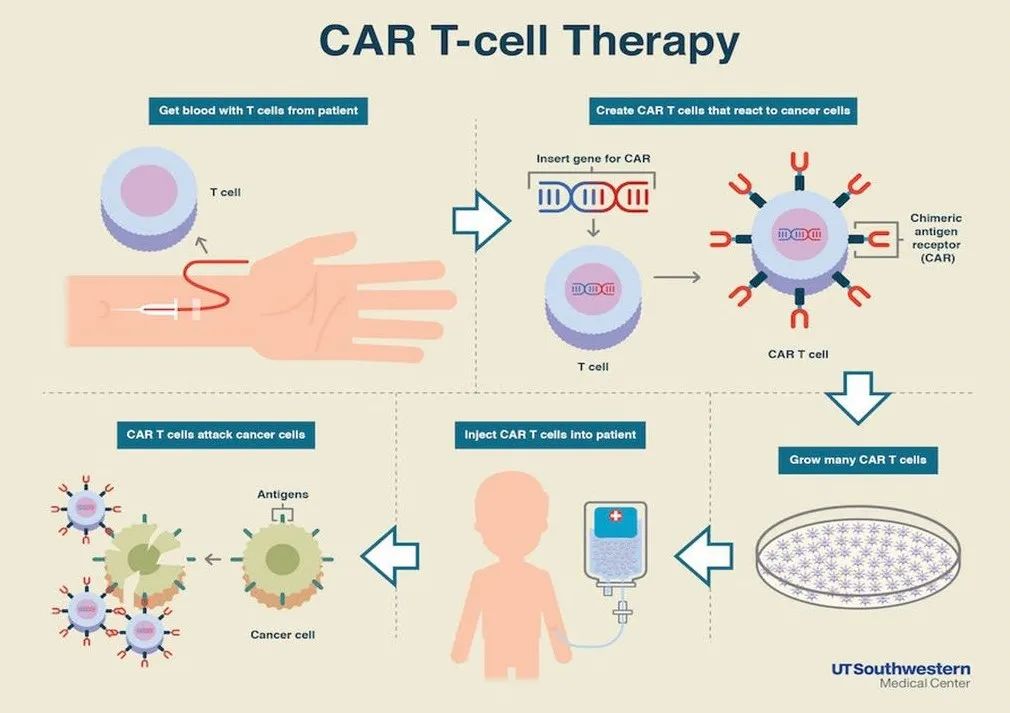
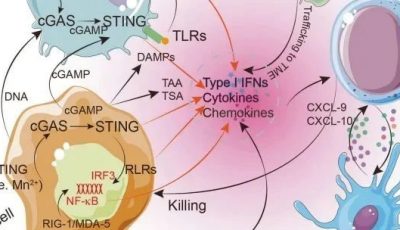
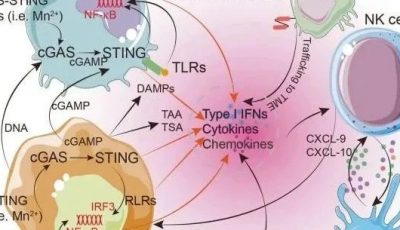
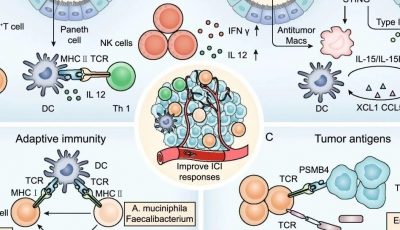
最新的癌症研究发现,基于免疫的治疗的成功需要能够识别突变靶点的T细胞受体谱系以及抗癌的细胞免疫反应(例如细胞毒性反应,Th1型免疫反应,见Fig4.1)。与此同时,基于T细胞的癌症免疫治疗正在快速发展,当前针对实体肿瘤的最成功的新型干预措施是依赖于癌症特异性T细胞活性及其在疾病部位的动员激活,即免疫检查点抑制剂疗法、嵌合抗原受体(CARS)和T细胞受体(TCR)修饰的T细胞也就是CAR-T疗法。抗原特异性肿瘤浸润性T淋巴细胞(TIL)的局部激活可以使细胞再循环,有力地杀死肿瘤细胞,减少肿瘤细胞数量,并在组织中协调抗肿瘤反应。
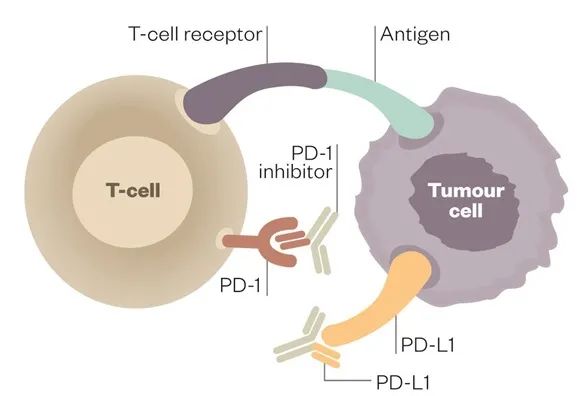
针对PD-1和CTLA-4的单克隆抗体为癌症治疗带来了革命性的变化,有迹象表明,它们有可能可以用于治疗病毒性肝炎、人类免疫缺陷病毒(HIV)感染、疟疾和结核病等慢性传染病。特别是抗 PD-1 疗法已被证明可以激活 CD8+ T 细胞,该细胞对与转移性黑色素瘤中的癌症进展相关的突变抗原(新抗原)具有特异性。在免疫治疗后患者血液中表现出新抗原特异性 T 细胞数量的持久增加,这提示了抗 PD-1 治疗的潜在机制。
4.2 肿瘤中的抗原加工和提呈
为了更好地理解推动抗原产生和对免疫系统“可见性”的动力机制,我们有必要了解一些关于抗原处理和呈递给免疫效应细胞的一些知识。抗原在自然条件下可以分为内源性的或外源性的;它们在细胞内经过生化处理,呈现给各种T细胞亚群、B细胞和自然杀伤(NK)细胞。与向免疫系统呈递抗原相关的基本分子被称为主要组织相容性复合体(MHC),在人类体内称为人类白细胞抗原(HLA)。MHC/HLA系统的功能是由Rolf Zinkernagel和Peter Doherty在20世纪70年代初发现并首次描述的,这项开创性的工作使他们在1996年获得了诺贝尔医学和生理学奖。
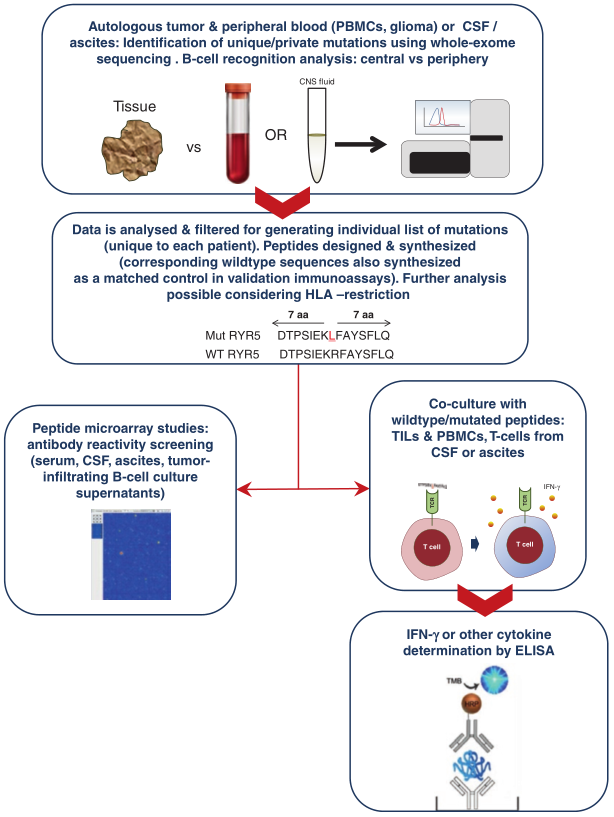
Fig.4.1 从转化细胞中识别突变表位的策略,CSF:脑脊液
抗原可以来自整个病原体,即细菌、病毒和寄生虫,也可以来自非突变或与转化细胞相关的突变蛋白。内在抗原,也被称为内源性抗原,经过处理后特定多肽(即表位)的形式呈现给免疫系统。这一途径被称为MHC/HLA-I类途径(以下称为“HLA I类途径”),该途径在诱导对病毒(在宿主细胞内合成的病毒成分)、细胞内细菌以及癌症相关抗原的免疫应答中起着至关重要的作用,而这些免疫应答依赖于免疫系统识别“自身”或“突变自身”抗原的能力。
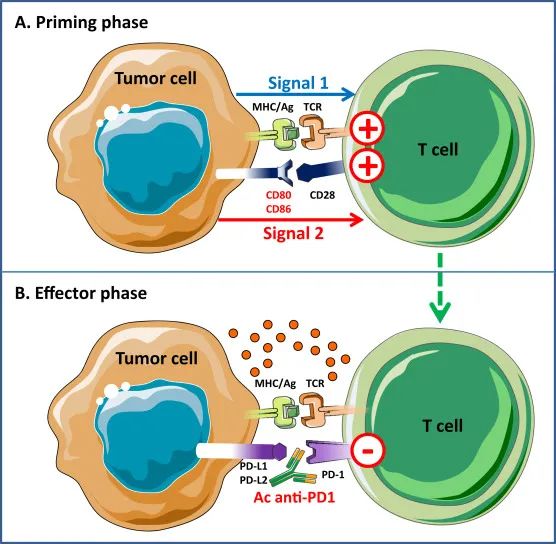
人体内的所有细胞(红细胞除外)都能通过HLA-I类途径处理和呈递抗原。在这个途径中,抗原的处理涉及到一个关键的步骤,即免疫蛋白酶体(发生在胞浆中)将变性(未折叠)的蛋白质结构切割成8到10个氨基酸长的小肽序列。蛋白酶体酶切蛋白质的氨基酸连接点决定了哪些肽或表位呈递给免疫细胞。在呈递给免疫系统后,CD8+T细胞可以识别HLA-I类分子呈递的表位,它可以通过增殖、产生细胞因子或产生能够杀死转化细胞的细胞毒性分子来做出反应。CD8+T细胞可以产生穿孔素、颗粒酶和粒溶素(使用CD107a诱导试验可以很容易地测量),或者通过-不是相互排斥的-干扰素-γ对转化细胞的反应。
如果(癌症)靶表位是用“反向免疫学策略”确定的,也就是说,表位是根据它们与MHC-I类或II类分子结合的预测能力来选择的,那么就不能非常确定地假设T细胞也会识别肿瘤细胞上自然加工和呈现的表位–而这种情况在20多年前就已经被描述过了:被证明多肽特异性的T细胞不能对肿瘤细胞上自然加工和呈递的多肽产生反应。导致这种现象的原因之一是,与负责激活和扩增抗原反应性 T 细胞的专职和非专职抗原呈递细胞相比,转化细胞中的特化或“扭曲”抗原加工和呈递机制可能不同 。另一种情况是,表位可能是通过翻译后修饰(如磷酸化)产生的,而这不能从野生型和/或突变蛋白的一级结构中预测出来。
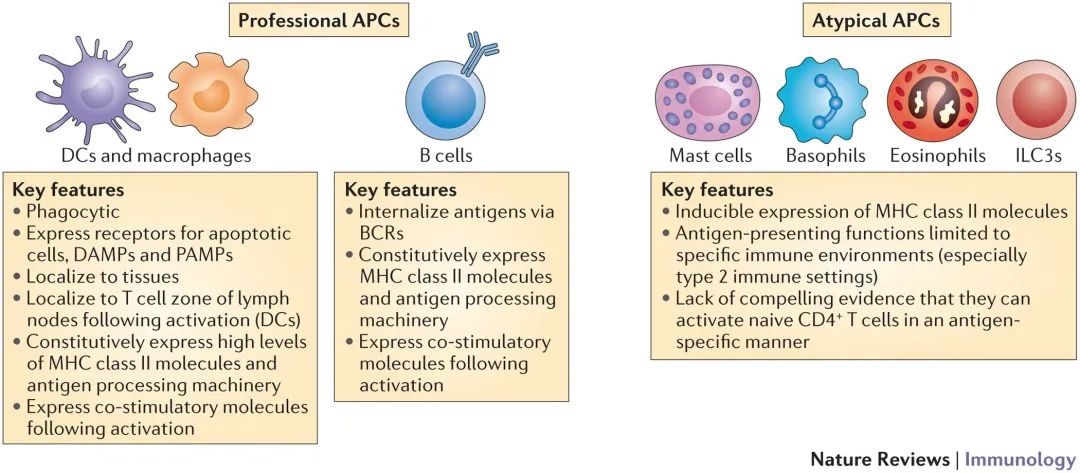
专业抗原提呈细胞或抗原提呈细胞(即树突状细胞、巨噬细胞),包括具有专业抗原提呈功能的B细胞,从外部环境中摄取的抗原通常被处理并通过HLA-II类途径提呈给免疫系统。整个的病原体以及因为抗体介导的机制、NK或CD8+T破坏癌细胞而产生的蛋白质,被抗原提呈细胞中称为吞噬小体的内吞囊泡主动摄取,之后溶酶体中包含的蛋白水解酶与吞噬小体融合以消化抗原,产生较小的肽序列,通常长度为13-17个氨基酸。这些表位被呈递给CD4+T细胞,也被称为辅助T细胞(Th),Th细胞可产生干扰素-γ、肿瘤坏死因子α(TNF-α)、白细胞介素(IL)-2(Th1细胞)、IL-4、IL-10(Th2细胞)等,在某些情况下也能通过产生IL-17(Th17细胞)协调免疫反应。细胞毒活性的产生并不完全归因于CD8+CTL;细胞毒性CD4+T细胞也被证明在癌症和病毒感染中介导生物相关的免疫反应。
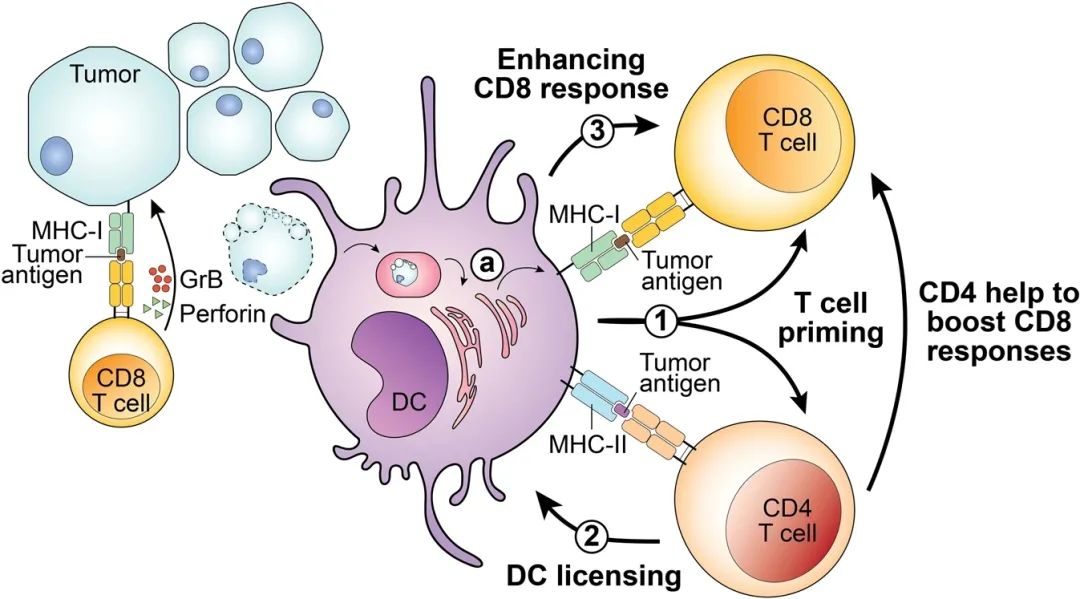
T细胞表面的T细胞受体(TCR)与辅助受体CD8或CD4结合到HLA表位复合体上,启动免疫突触。HLA限制T细胞和肿瘤细胞之间的相互作用,并与组织微环境或血液中可用的TCR相匹配,这些过程决定了免疫反应的性质和强度。
HLA抗原表位的限制性和免疫交叉反应性在决定免疫应答的性质中起着不可或缺的作用。例如,人类白细胞抗原-DQ变异体与某些传染病的易感性增加有关;人类白细胞抗原-DQ基因β57亚单位的突变可能导致肺部疾病的形成。有趣的是,HLA-DQ等位基因突变被归因于1型糖尿病(T1 DM)的易感性。虽然HLA-DQ在美洲和欧洲的高加索人中非常普遍,但东亚人和非洲人表达这些等位基因的可能性要小得多。
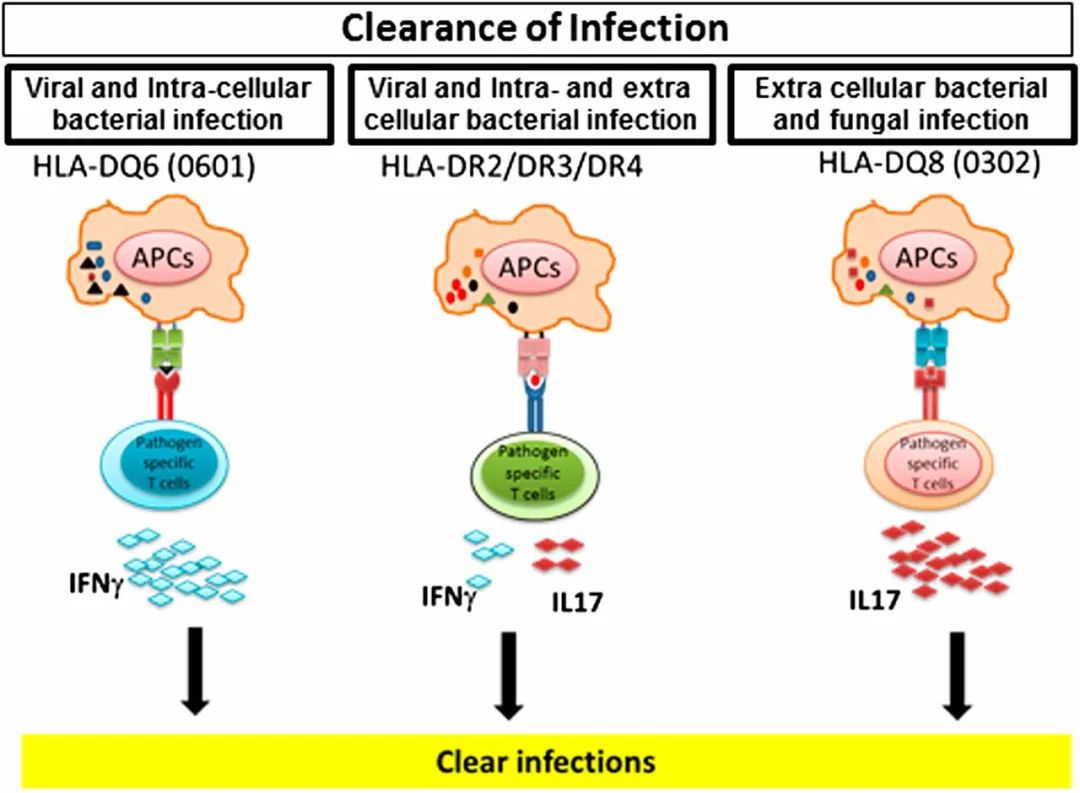
事实上,个别的HLA等位基因也可能有利于某些免疫识别特征,如HLA-DQ0602有利于IL-17的产生,而不依赖于结合肽,正如多发性硬化症转基因小鼠模型所显示的那样。这种以IL-17为中心的反应是一把双刃剑;它可能更有效地遏制某些细菌感染,也有可能有利于吸引免疫细胞到肿瘤部位,而长期暴露于IL-17可能反而会促进恶性转化。因此,疫苗接种后免疫反应的性质、质量和数量似乎在很大程度上取决于个人的HLA谱,HLA谱塑造了随后机体的细胞免疫反应的质量和数量,包括感染风险的增加或降低。自身免疫反应或向T细胞呈递(Neo)表位的能力取决于MHC-肽复合物和响应的TCR谱系施加的限制。例如,即使新表位是在恶性转化过程中产生的,但如果它们没有经过处理并最终与相应的HLA分子复合并呈现给反应的T细胞,它们可能不会被细胞免疫系统识别。
Th1/Th2应答以及MHC的限制性
大多数研究使用IFN-γ作为T细胞对野生型癌细胞和突变表位作出反应的测量指标。Th2反应以IL-4、IL-5的表达为标志,同样也可以表达IL-13,这些细胞因子可能是原发的Th2反应产生的,也可能是Th1T细胞转变为的产生Th2细胞因子的T细胞所产生的,这种Th1到Th2的转变可能是由突变事件产生的部分激动肽的结果。
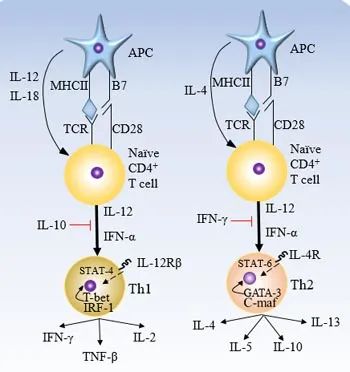
Th2型T细胞反应本身可能并不意味着抑制性和潜在的“耐受性”免疫反应;最近的报道表明Th2型免疫反应也可能介导产生临床相关的抗肿瘤免疫反应。在临床前模型中,抗原特异性Th2细胞在没有CD8T细胞的帮助下根除了骨髓瘤,导致了肿瘤部位的大范围炎症。研究表明,Th2介导的肿瘤破坏与原位产生的IL-1、TNF-α(Th1)和Th2细胞因子(IL-4、IL-5、IL-13)有关,而被动转移的Th2细胞能够产生持久的细胞抗肿瘤免疫反应。
在Th2介导的免疫应答中,CD8非依赖性和抗原特异性T细胞是嗜酸性粒细胞趋化因子(Eotaxin)和STAT6依赖性的T细胞。而人类癌症中的Th2浸润还没有得到广泛的研究,一些研究甚至表明使用Th2型细胞因子会产生更好的免疫结果。Th2反应在识别突变表位方面的实质目前还没有得到很好的研究。CD4-Th2反应之间的关联也可能受益于T细胞与限制性的MHC II类分子的更密切的关联。例如,以前的研究报道,在DPB1*0401/0402阳性的卵巢癌患者中,Th1/Th2 CD4+T细胞对NY-ESO-1有反应。而更多的信息是关于针对来自感染性病原体的相对较少变异(与HLA-DR相比)的HLA-DP分子所呈现的多肽的细胞免疫反应的性质,例如乙型肝炎或MHC II类分子,它们使人类容易患上某些自身免疫性疾病(例如麸质相关性结肠炎)。某些MHC等位基因与某些细胞因子产生模式(IL-17、Th1、Th2)相关的变异表位的影响到目前为止尚不清楚。Table.4.1提供了从胶质瘤患者的TIL中识别的野生型和突变型靶表位的概述,表明针对突变型表位的单个患者的TCR谱系中存在Th2反应。
新抗原的加工和呈递可能会产生突变表位(新表位),这些表位既是共享的,也是患者特有的。这当然取决于突变的位置,即可能破坏自然发生的裂解位点的点突变,以及突变本身的性质,即点突变、染色体缺失和过早终止密码子。使用从代表20种不同癌症类型的患者的肿瘤和非肿瘤组织中提取的DNA,对HLA-I类途径中的体细胞突变进行综合分析,发现HLA-I类分子的N端发生功能丧失突变的可能性很高,它阻止了多肽-HLA复合物向细胞表面的运输。此外,在所有被测试的癌症中,发现最常见的突变发生在HLAI类分子的CD8-TCR 3区域,这一突变是在免疫突触过程中T细胞上的α共同受体结合所必需的,通过该突变从而激活CD8-TCR复合物。
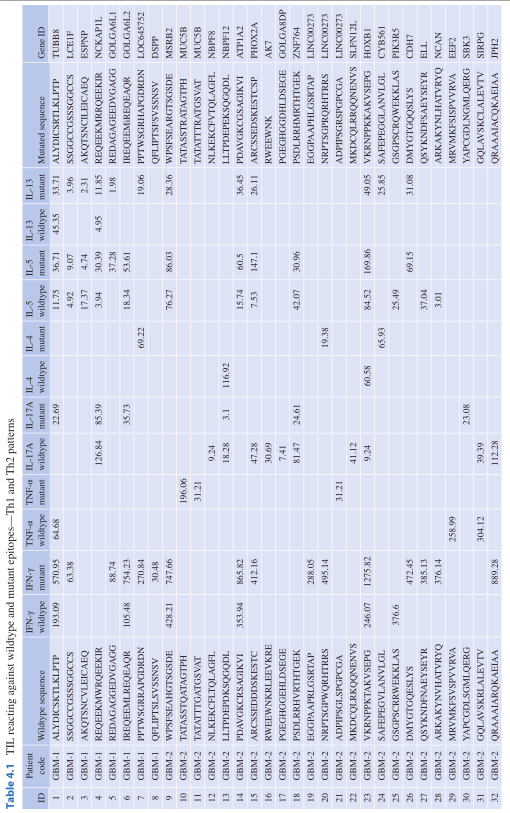

Table.4.1 针对野生型和突变型表位的TIL-Th1和Th2模式
4.3 肿瘤抗原和表位:从发现到治疗应用
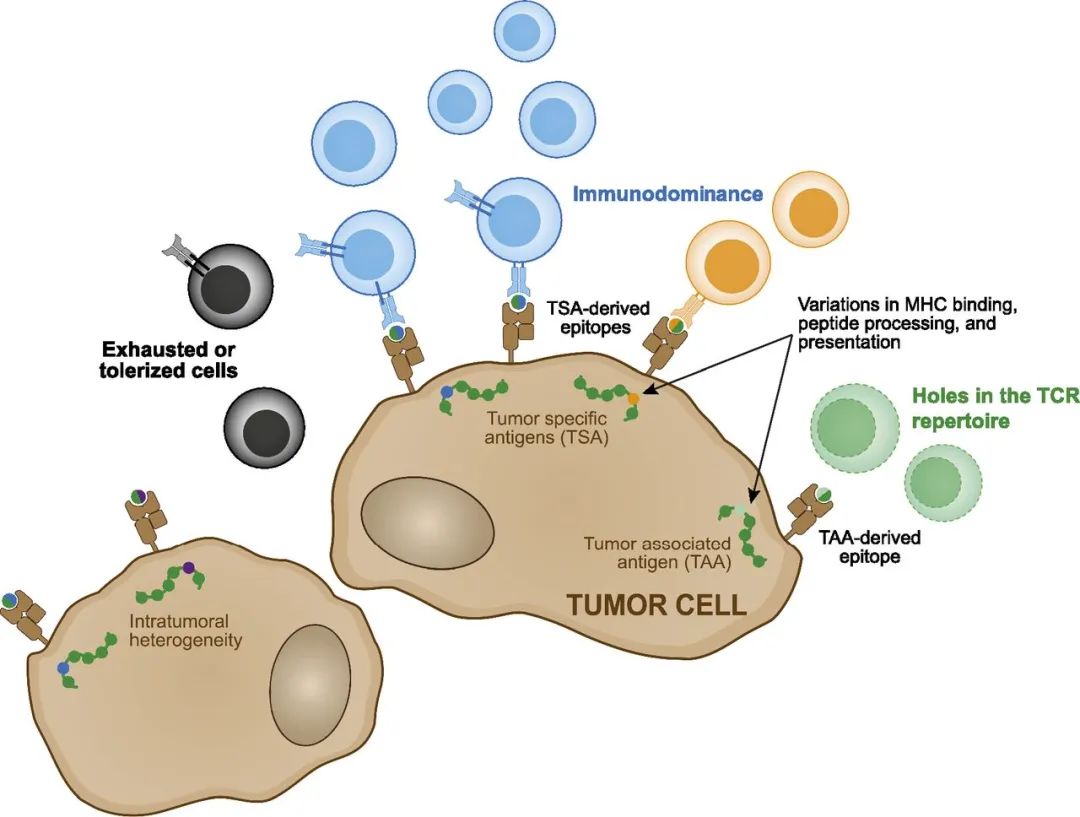
人类癌症,特别是黑色素瘤小鼠模型的临床前研究,在肿瘤排斥的背景下,提供了对癌症抗原发现和功能特征的第一次实验观察。Thierry Boon及其同事在20世纪80年代末曾证明,在小鼠P815肿瘤细胞中异源表达的肿瘤抗原P19A(从携带甲基胆蒽诱导的肉瘤的DBA/2小鼠中分离出来)包含一个HLA-I类表位(位于一个含有点突变的13聚体序列中),能够引发有效的CTL反应和靶细胞的裂解。
由欧洲和美国的研究小组带头,首先使用从人类黑色素瘤病变中提取的肿瘤组织,在人类癌症环境中进行表位挖掘。比利时布鲁塞尔路德维希研究所(Ludwig Institute)的蒂埃里·布恩(Thierry Boon)、皮埃尔·库利(Pierre Coulie)和他的同事们于1991年发现了第一个肿瘤相关抗原(TAA),被一株自体CTL株识别,并诱导表达MAGE-1DNA的肿瘤细胞株裂解,并受到HLA-A1的限制。此前他们使用来自一位患有转移性疾病的匿名患者MZ2的黑色素瘤细胞株体外鉴定了CTL反应。这种TAA最初被注释为MZ2-E,后来更名为黑色素瘤相关抗原1(MAGE-1,癌症睾丸抗原)。通过对同一患者的细胞系的进一步研究,他们发现了MZ2F,也就是今天所知的G抗原1(GAGE-1)。当时正在进行的大部分工作集中在发现新的免疫原性HLA -I类限制性抗原,这些抗原介导了患者黑色素瘤细胞的CTL反应和溶解,人们有浓厚的兴趣首先了解其机制并开发基于免疫的干预措施:MAGE-3.A1肽免疫患者中的Melan-A(HLA-A2+表位)和MAGE-3(HLA-A1+表位)特异性CTL反应。
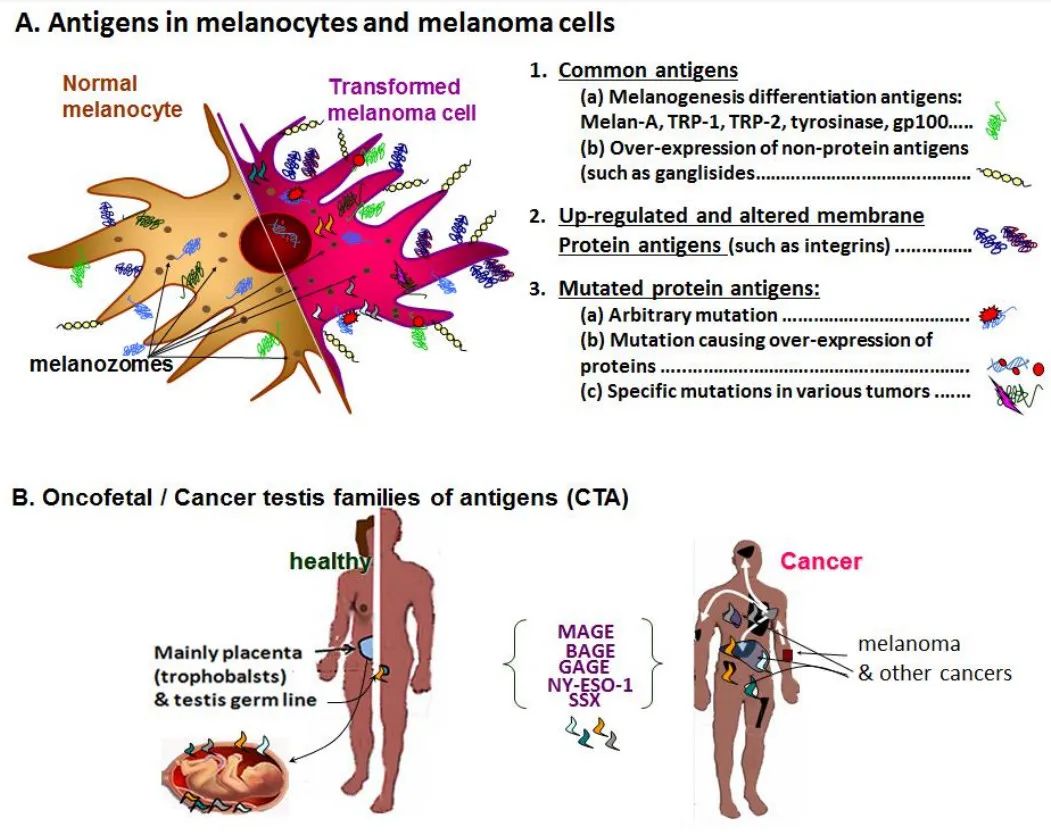
欧洲和美国的研究人员同时发现了另一种重要的癌症抗原—癌症睾丸抗原NY-ESO-1,它是用从人食管鳞癌细胞制备的cDNA对表达cDNA文库(SEREX)进行血清学分析(表明存在抗体反应)而发现的。Elke Jäger和他的同事(法兰克福)后来发现NY-ESO-1含有具有生物学功能的CD8+(HLA-A2/B51)和CD4+(HLA-DRB*1)T细胞表位,这些发现是基于对人黑色素瘤细胞和转染T2细胞的精液研究。上述T2细胞在与抗原处理(TAP)相关的转运体中存在缺陷,从而抑制它们呈递内源性胞浆多肽(除了一些装载在HLA-A2分子上的前导肽序列),但允许引入外源添加的HLA-I类表位用于CTL识别试验。
史蒂文·罗森伯格(Steven Rosenberg)和他在美国国家癌症研究所(NCI,National Institutes of Health(NIH),Bethesda,MD)外科分院的同事对人类黑色素瘤抗原的发现做出了关键贡献,特别是发现了一些诱导TIL之间的反应性的抗原:1995年发现由HLA-A31分子限制的酪氨酸相关蛋白1(Trp-1)或gp75;HLA-A31限制性Trp-2肽LLPGGRPYR,它是TIL输入转移性黑色素瘤患者的主要靶点,后来用于显示疾病消退程度;由HLA-A31和HLA-A33限制的Trp-1和Trp-2(TRP197-205)表位;由HLA-DR1限制的磷酸丙糖异构酶产生的突变表位,由CD4+TIL和细胞分裂周期蛋白27同系物(CDC27)识别。总体而言,这些早期的研究努力(从20世纪80年代末到21世纪初的15年间)为未来研究提供了一个极好的基础,进一步推动靶向癌症免疫治疗领域的扩大。
随后,一系列其他分子被确认与转化细胞有关。例如,间皮蛋白被发现是几种重要的实体癌症的标志物,即间皮瘤、卵巢癌、胰腺导管腺癌,而这些都是通过血清学(一种识别人间皮蛋白的小鼠‘KI抗体’)和基因分析发现的。接着进一步探讨该与转化细胞相关的分子在卵巢癌、间皮瘤和鳞癌中的临床意义,并结合患者血清中可测的间皮素和抗体反应,来表明间皮素具有免疫原性并被指定为癌症抗原。这种基于K1抗体间硫蛋白结合区的实验性免疫毒素是最早被尝试于基于靶向免疫的干预措施之一,而这些临床前研究都是在人类肿瘤异种移植的小鼠模型上进行的。
20世纪90年代后期实施的工作将更多的重点放在研究人类癌细胞中的突变蛋白,以及发现由HLA限制性元件呈现的具有生物学和临床意义的突变抗原决定簇(新表位)在临床免疫治疗的可能性。一个早期的例子是黑色素瘤无处不在的突变1蛋白(MUM-1,最初命名为LB33-B,以黑色素瘤患者LB33的名字命名)上的新表位,该表位受HLA-B*44*02等位基因的限制。该9聚体新表位是在对自体黑色素瘤细胞系LB33-MEL.A-1进行体外细胞毒性实验后确定的;而在野生型多肽序列中没有发现有与其相同的细胞杀伤活性的表位。突变细胞的细胞周期受细胞周期素依赖性激酶4(CDK4R24C)蛋白的10聚体新表位(氨基酸23-32)受HLA-A*0201限制,在转染CDK4R24C cDNA的T2细胞作用下,可通过自体CTL介导细胞杀伤活性,且呈剂量依赖性。而HLA-B*3503限制性半胱氨酸天冬氨酸蛋白酶8(Caspase8)突变肽对自体头颈癌细胞和转染肿瘤cDNA的B细胞具有较强的杀伤活性。这些研究进一步扩大了从人癌细胞中挖掘新表位的研究领域。
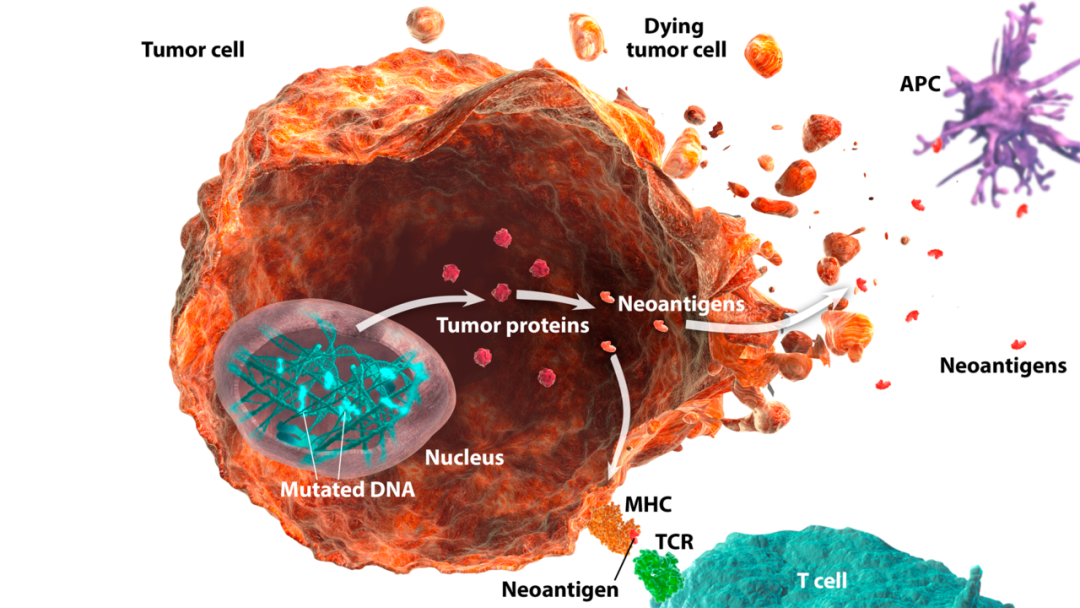
对代表30种不同人类癌症的临床肿瘤样本的整个基因组和外周DNA的高通量分析揭示了每种癌症类型中独特的突变负荷,以及这些癌症特有的突变特征。虽然这提供了人类癌症中突变负荷的总体图谱,但每个患者的突变特征是不同的-因此产生了一份私人突变特征的“概要”,这些突变特征不仅涉及驱动和维持恶性转化,而且涉及免疫效应细胞的激活和扩增。
V-Ki-ras2Kirsten大鼠肉瘤病毒癌基因是其同源基因的突变形式,简称KRAS,是一种公认的新抗原,与胰腺癌、结直肠癌和肺癌的发病有关。天然KRAS是1982年在人类肺腺癌基因测序后发现的,是一种参与细胞信号转导的鸟嘌呤三磷酸酶;然而其12、13和16位的突变与肿瘤的发生有关,从而使其成为人类的原癌基因。
美国国立卫生研究院(Bethesda,MD)外科分部的Steven Rosenberg和他的同事最近开发了一种尖端方法,为个别患者筛选可以识别新表位的新表位特异性T细胞反应。这种方法被称为“串联微基因(TMG)”方法,它首先需要从患者肿瘤组织样本中提取的基因组DNA的全外显子测序数据。而测序数据会产生病人基因编码DNA中包含的所有非体细胞突变,这样就可以构建一个患者“私人”突变的个人文库,而这些突变可能编码新表位。然后,通过人工把这些短基因序列构建在一起,并插入到表达质粒中,该表达质粒随后被转染到慢病毒载体中,用于感染APC(来自患者的树突状细胞)。然后,将来自同一患者的自体TIL与携带TMG的树突状细胞共培养,以诱导免疫反应。以TIL产生干扰素-γ为代表的阳性反应将标志着共培养的DC含有TMG,该TMG包括一个新的表位编码序列,该序列会被自然处理并呈递给免疫系统。
基于突变表位确实会被处理并呈递给T细胞的前提条件,不同种的会对突变产生响应的T细胞群体可能会对该突变产生不同的、互不排斥的影响:(1)新招募的T细胞将特异性识别突变序列 (2)野生型和突变型表位序列之间存在潜在的T细胞受体(TCR)交叉反应(如果两者都有突变表位,则野生型和突变型表位会直接被处理并呈递给T细胞)。是否已经存在可以识别野生型表位的细胞和在恶性转化后识别突变靶点的T细胞群体是否已经扩大,这两个问题在生物学上是相关的,因为这些情况可能导致不同的TCR触发,并随后导致不同的T细胞效应器功能。
突变表位作为对抗癌细胞的“生物手术刀”,通过招募识别突变表位序列的T细胞来增强免疫效应功能,是一种临床上有吸引力的尝试,目的是改进和扩大针对仅存在于恶性肿瘤中的突变的TCR谱系,因此将代表“理想的”癌症相关抗原-一种只针对癌细胞而不会伤害未转化细胞的“生物手术刀”。如何利用突变表位的这一特点呢?一种方法是在不与MHC限制性分子相互作用但与TCR接触残基相互作用的残基上修饰多肽——这些变体被称为“异位”类似物,能够触发具有不同T细胞效应功能的TCR,例如细胞毒性、细胞因子产生的质量和数量以及细胞增殖。这种方法已经被用在诱导T细胞对野生型多肽(例如对p53)产生反应上。如果表位发生单一突变,那可能会产生由肿瘤细胞自然加工并呈递给TCR的多肽。

如果针对野生型靶点的T细胞反应也能够对由相同的HLA限制元件呈现的相应突变靶点表位做出反应,那么可能会发生以下情况:虽然每个单独TCR的单一配体特异性似乎很少见,但是TCR的识别已被证明是灵活的,并由突变的肽配体诱导,分为拮抗剂、部分激动剂和超级激动剂。
多肽靶标内的突变可以诱导TCR/CD3复合物的差异磷酸化,并具有不同的下游信号通路配置。因此,如果野生型多肽也能被识别,突变可能就会因为配体没有被处理和呈递,而导致T细胞识别功能丧失。另一方面,也可能出现T细胞配体可以被很好地处理和呈递,但T细胞信号可能由于细胞无能而被清除的现象。部分激动剂,通过诱导单个氨基酸残基,即使在野生型多肽与表位的相似剂量范围内,仍然能够刺激针对野生型多肽的T细胞反应:当带有野生型TCR配体的T细胞与增殖、细胞因子产生和细胞毒性反应时,多肽变体可以在没有T细胞增殖的情况下诱导细胞毒性和或细胞因子产生。
值得注意的是,类似的观察结果可能适用于针对突变表位的TCR,该表位会与定性和定量不同的T细胞反应模式发生反应。单一氨基酸交换也可能使T细胞克隆产生Th1细胞因子转变为产生Th2细胞因子,或者导致T细胞克隆停止产生细胞因子,但如病毒病原体所显示的那样,T细胞仍具有强烈的细胞毒性T细胞反应。靶点的突变与不同的信号事件相关,也可能在T细胞应对野生型和突变靶点时的分化状态的调节起关键作用,这一点在胸腺细胞分化和成熟的临床前模型中有说明。TCR的不同触发与T细胞的成熟和分化有关,而T细胞的成熟和分化质量对于长期免疫记忆、接触(肿瘤)组织以及T细胞治疗的临床疗效都是十分重要的。
临床试验证明,针对TAA(一种有机化合物)的免疫细胞会被动转移,同时发现T细胞产物的转移会导致中央记忆T细胞的产生。控制T细胞记忆的生化信号不仅依赖于细胞因子环境,还依赖于TCR-CD8链复合体传递的信号强度。包括T细胞记忆在内的T细胞反应的质量和数量由ζ肽-TCR相互作用的强度显著决定,这一现象可能在一定程度上与CD8+T细胞有关。现在已经提出了一个降低电位模型,用于测量目标表位传递给相应TCR的信号强度,该TCR决定T细胞是否有可能进入T细胞存储池。这个“信号强度”模型需要考虑T细胞产生的细胞因子和与T细胞进入记忆T细胞池的多样性相关的促炎信号。通常,较弱的TCR信号就足以将T单元移动到存储器T单元编程中。TCR信号的长度(即缩短TCR刺激)也将决定T细胞是否进入记忆T细胞池,这两种决定方式是不相互排斥的。并且观察到由TCR依赖的NFKB信号介导的多肽中的点突变影响了与TCR的接触,这也损害了CD8+T细胞的记忆发育。这可能部分解释了为什么针对相同(突变)肿瘤表位的T细胞克隆存在异质分化状态(见Fig.4.2)。
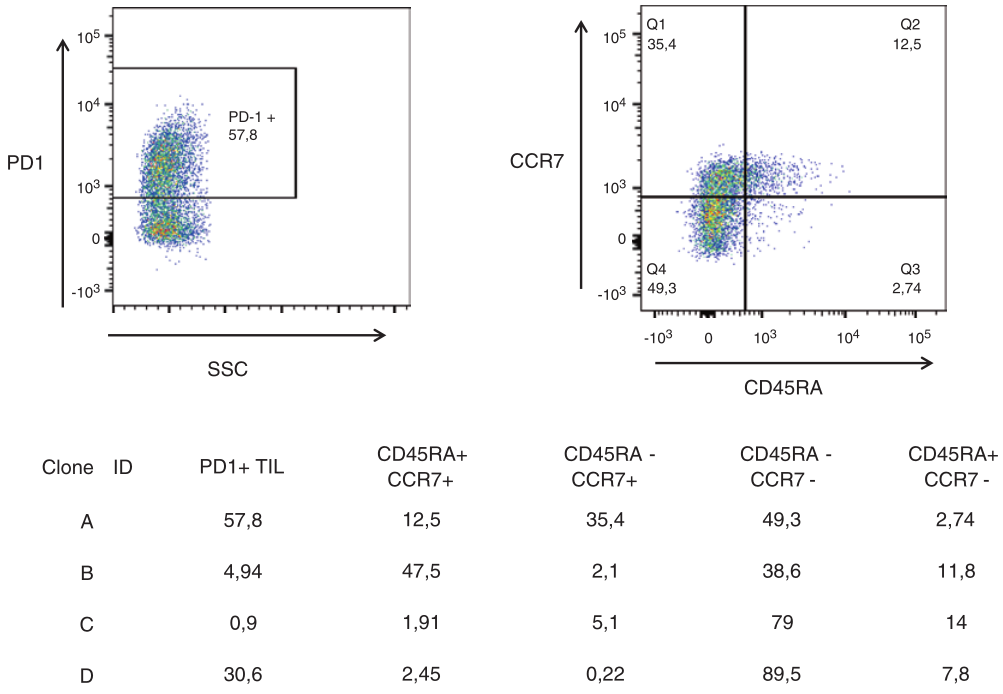
Fig 4.2 不同的T细胞克隆(A:VB5.1,B-D:VB9)识别自体胰腺癌细胞上自然加工和呈现的(突变)靶点。值得注意的是,肿瘤导向的T细胞克隆表达不同的T细胞归巢和分化标志物,由CD45RA和CCR7的表达定义,即CD45RA+CCR7+T细胞是前体T细胞,CD45RA-CCR7+中央记忆T细胞,CD45RA-CCR7-T细胞记忆效应T细胞和CD45RA+CCR7-T细胞代表终末分化效应T细胞。其中,大多数来自克隆B的细胞位于中央记忆T细胞亚群中,该亚群已被证明与癌症细胞治疗中增强的反应性有关。
由于对抗原的非经典识别,γ-δ(γδ)T细胞在癌症中的作用也被重新探讨。γδT细胞识别非肽结构,即由CD1d分子呈现的磷酸化抗原,如真核类异戊二烯(甲羟戊酸)途径的衍生物。Vγ9Vδ2(Vδ2+)的γδT细胞亚群,存在于外周血中。它们还被描述为表达CXCR3的表面标记,这对组织穿透至关重要,这是机体接触转化细胞或组织驻留病原体的重要特征。Vδ2+γδT细胞的一个有趣特征是表达CD16共受体,该受体除了能与杀伤受体NKG2A结合外,还能与肿瘤细胞上存在的FcγRIII结合。因此,像NK细胞一样,γδT细胞也可以协调抗体依赖性细胞毒作用,这一结论与几种基于单克隆抗体的抗癌药物,如利妥昔单抗、曲妥珠单抗、奥曲单抗和阿仑单抗的治疗活性有关。异戊二烯焦磷酸酯(IPP)是异戊二烯途径的中间产物,可以被Vγ9Vδ2 T细胞强烈识别,如唑来膦酸处理的人类癌细胞所显示的那样。唑来膦酸可诱导肿瘤细胞内异丙肾上腺素积聚,从而刺激Vγ9Vδ2T细胞的活性,进而促进干扰素-γ和细胞毒分子的产生。在IL-2和/或IL-15调节的存在下,这一效应可以进一步增强。虽然改变/突变形式的IPP尚未被报道,但γδT细胞在靶向细胞治疗中的意义仍需进一步探讨。与此同时,也有报道称γδT细胞能够识别和杀伤癌细胞上过表达的人热休克蛋白60/70,这表明人类识别肿瘤抗原的T细胞谱系是相当丰富的。
— THE END —
来源 | Oncoimmunology: A Practical Guide
for Cancer Immunotherapy
作者 | Martin Rao, Liu Zhenjiang, Qingda Meng, Georges Sinclair,
Ernest Dodoo, and Markus Maeurer
翻译 | 郑梦怡 徐曼斯
编辑排版 | 钟雪莹
初审 | 宋乐 陈秋谷
终审 | 肖建勇 王坤
(转载请保留此部分内容)
▉ 往期精彩内容推荐
基础免疫学系列
肿瘤免疫学系列
肿瘤免疫学-第3章:CD8+ T细胞在免疫治疗、放疗和化疗中的作用(下)
肿瘤免疫学-第3章:CD8+ T细胞在免疫治疗、放疗和化疗中的作用(上)
生信入门学习笔记系列
基础实验技术系列
Annexin V/PI染色检测细胞凋亡-OncoLab实验室
仪器操作系列
蔡司LSM800激光共聚焦显微镜操作视频-中西医结合基础研究中心
IN Cell Analyzer2200高内涵细胞成像分析系统操作教程-中西医结合基础研究中心
文献分享系列
年度巨献:神刊CA发布最新癌症数据:全球1000万人死于癌症,中国占比超3成
HLA-I分子加工和递呈抗原功能受损是免疫检查点抑制剂耐药的重要原因
作图投稿选刊系列
GraphPad+AI制作可发表级别的柱状图-OncoLab实验室
PubMed中如何按影响因子筛选检索结果并显示影响因子及分区?
如何使用EndNote软件插入参考文献(附6813种杂志参考文献格式文件)
中医中药系列

关注本号~

加入读者交流群~
本篇文章来源于微信公众号:OncoLab