4.4 感觉信息处理在大脑皮层达到顶峰
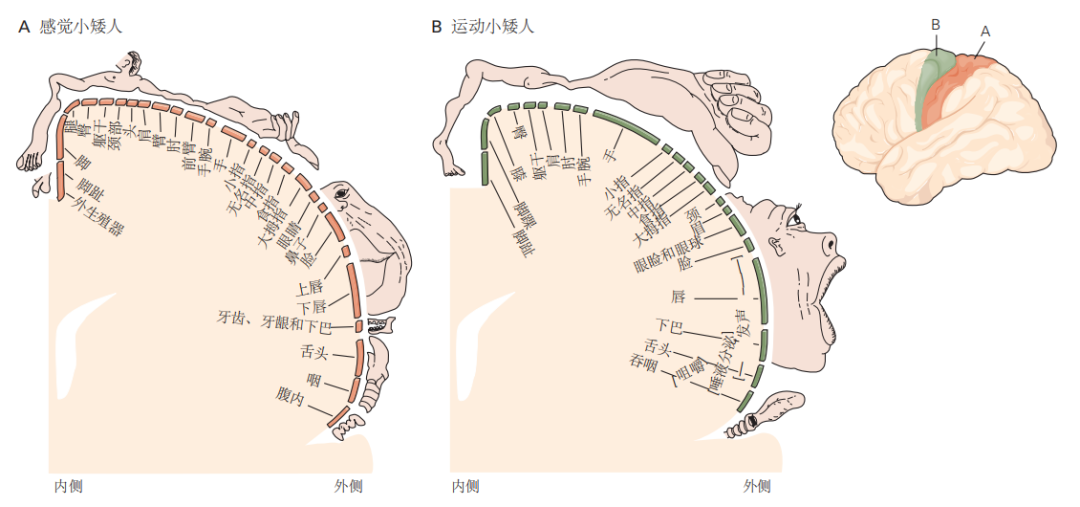
图 4.4.1:小矮人图示展示了专门用于控制身体各部位的感觉和运动神经支配的皮层区域的相对大小。整个身体表面在皮层中按有序的体感输入图表示。
A. 皮层区域的大小并不与身体部位的体积成正比,而是反映了该部位感受器的密度。因此,嘴唇和手的感觉输入占据的皮层区域比肘部更多。
B. 运动皮层的输出也以类似的方式组织。专用于身体某个部位的皮层表面积与该部位的运动控制精细度相关。因此,在人类中,大部分运动皮层用于控制手指肌肉和与语言相关的肌肉。
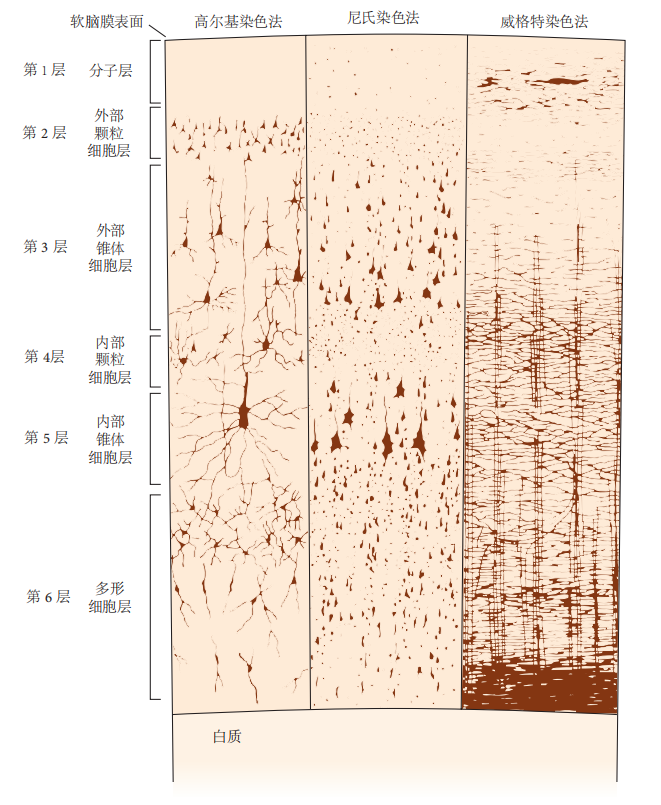
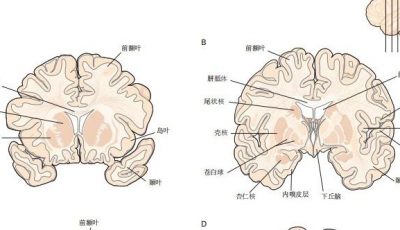
图 4.4.2:新皮层的神经元排列成不同层次。根据所使用的染色法,皮层外观有所不同:高尔基染色法(左)揭示神经元的细胞体、轴突和树突;尼氏染色法(中)显示细胞体和近端树突;威格特染色法(右)揭示髓鞘纤维的模式。
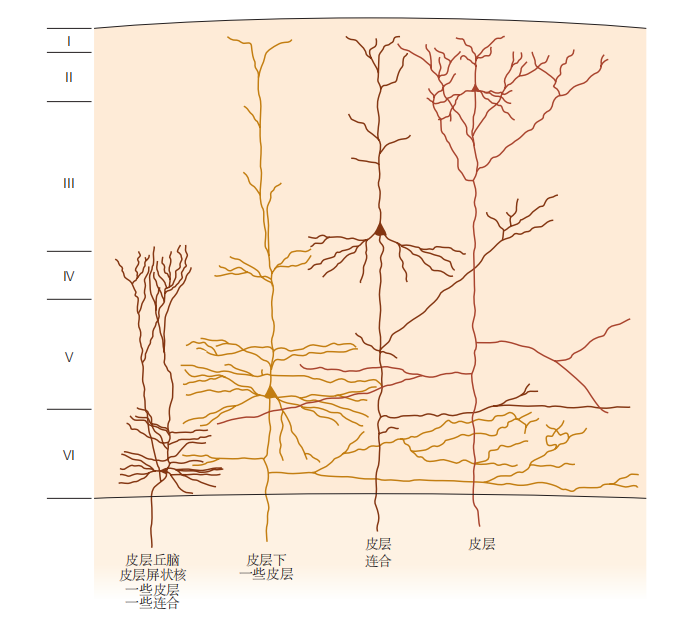
图 4.4.3:新皮层的不同层次神经元投射到大脑的不同部分。新皮层的联合连接主要由第2层和第3层的神经元介导,而投射到皮层下区域的神经元主要来自第5层和第6层。
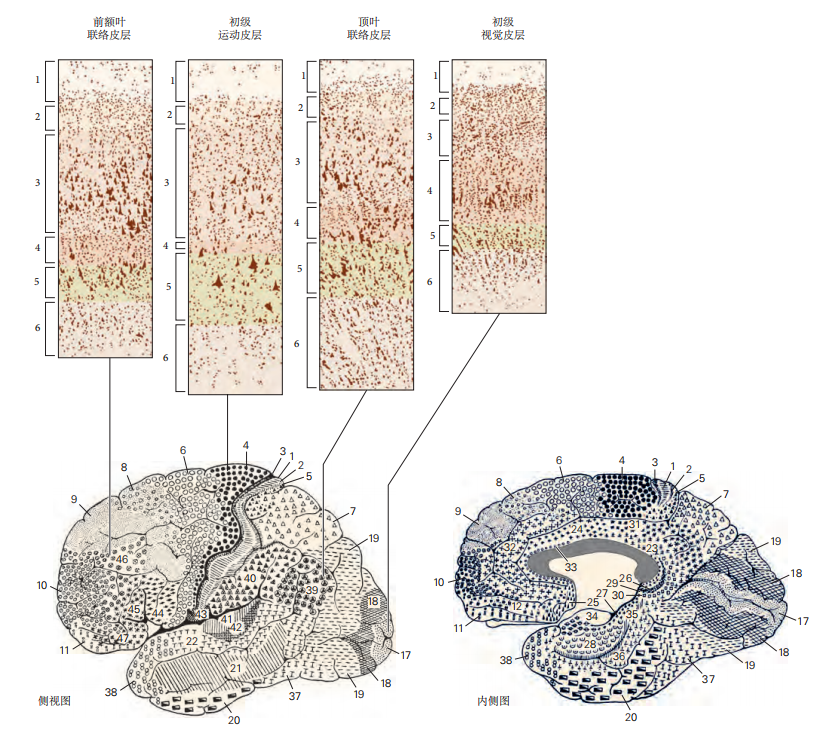
图 4.4.4:新皮层的细胞层在各个皮层区域有不同的表现。感觉区域(如初级视觉皮层)具有突出的第四层,而运动区域(如初级运动皮层)则表现为薄弱的第四层,但具有显著的输出层(如第五层)。布罗德曼的皮层细分是基于细胞结构的经典分析,至今仍被广泛参考。
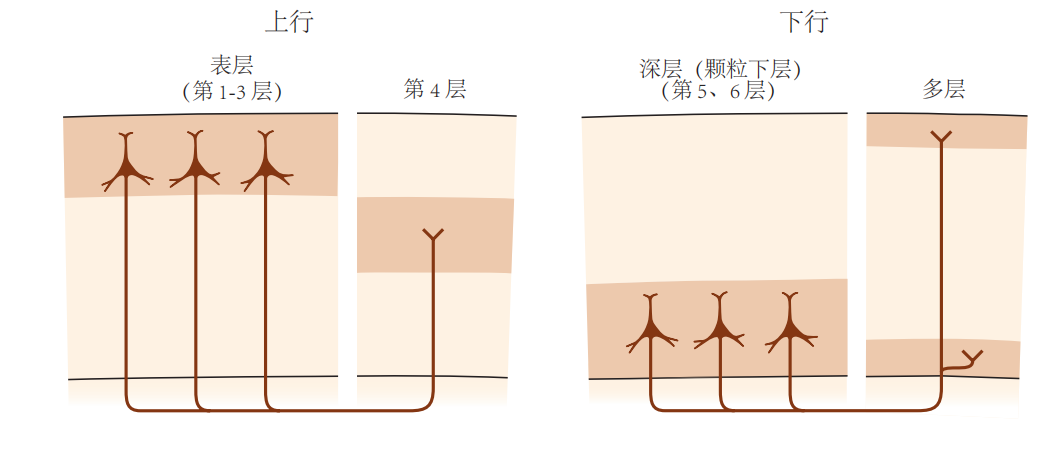
图 4.4.5:上行皮层通路和下行皮层通路的区别在于它们在皮层内的起源和终止位置。上行通路(或前馈通路)通常起源于皮层的表层,并总是终止于第4层;而下行通路(或反馈路径)则起源于皮层的深层,并终止于第1层和第6层。
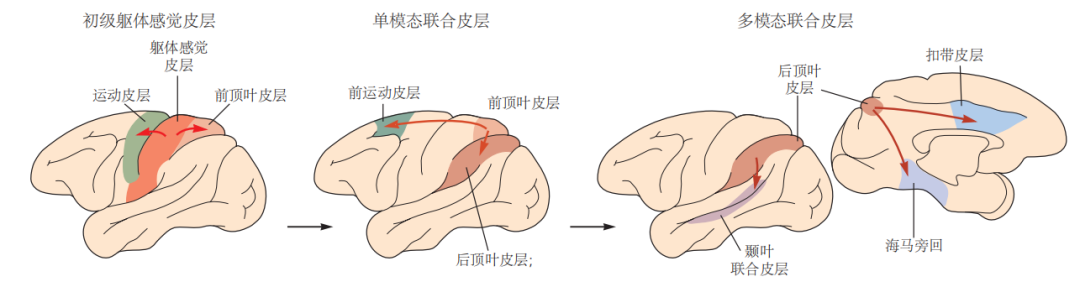
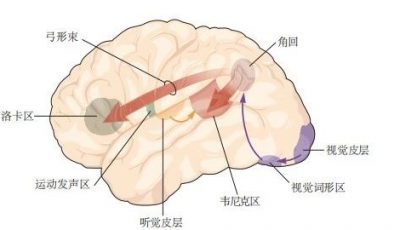
图 4.4.6:大脑皮层中的感觉信息处理从初级感觉区开始,经过单模态联合区,最终在多模态联合区进一步整合。感觉系统还与运动皮层相互通信。例如,初级躯体感觉皮层投射到额叶运动皮层和顶叶的躯体感觉联合皮层,而躯体感觉联合皮层则反向投射到高阶躯体感觉联合区和前运动皮层。来自不同感觉系统的信息汇聚在多模态联合区域,其中包括海马旁回、颞叶联合皮层和扣带皮层。
4.5 自主运动由皮层和脊髓之间的直接连接介导
4.6 大脑中的调节系统影响动机、情绪和记忆
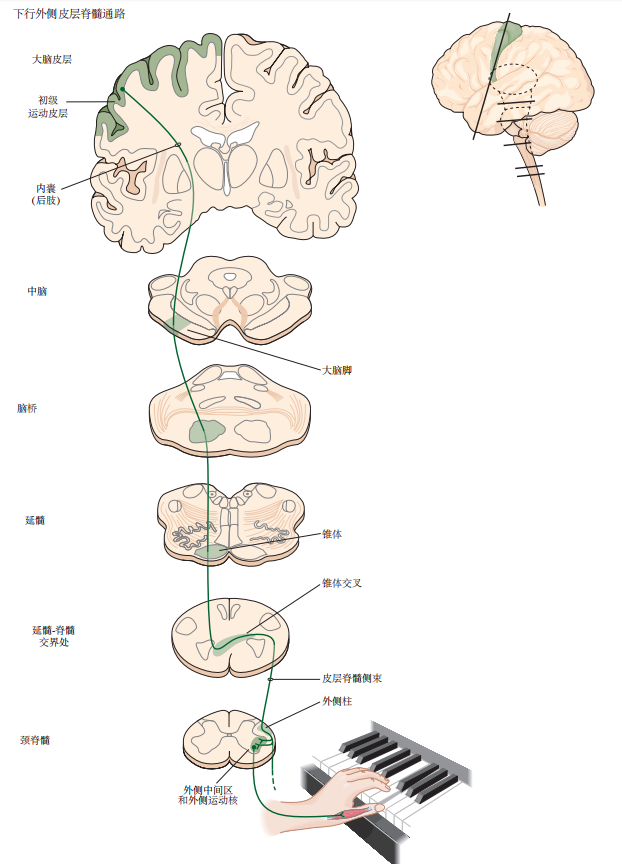
图 4.5.1:皮层脊髓束中的大量纤维起源于初级运动皮层,终止于脊髓的腹角。相同的轴突在其投射的不同阶段分别形成内囊、大脑脚、髓质金字塔和外侧皮层脊髓束的一部分。
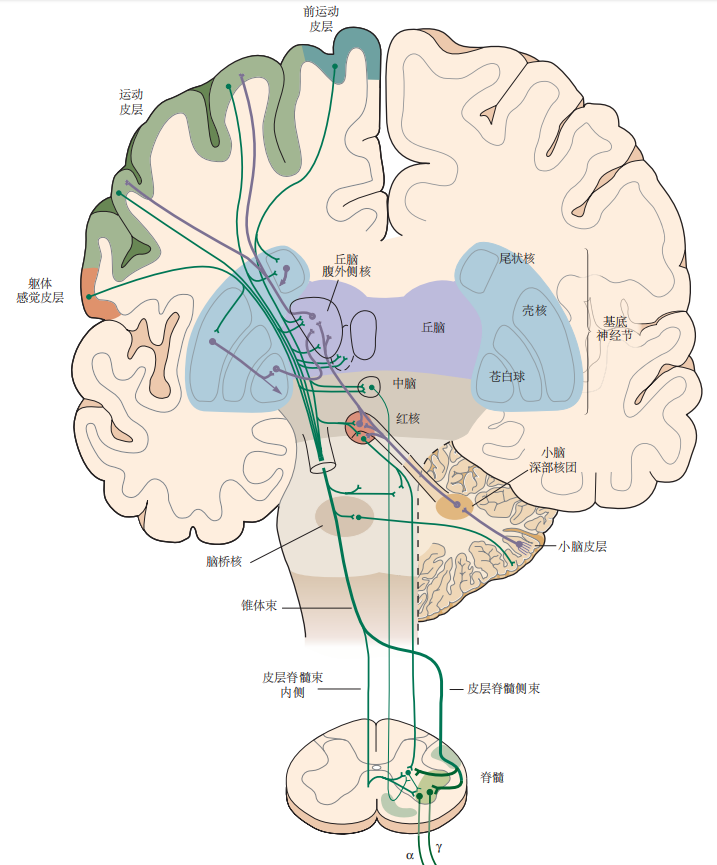
图 4.5.2:自主运动需要协调运动系统的所有组成部分。主要组件包括运动皮层、基底神经节、丘脑、中脑、小脑和脊髓。主要的下行投射以绿色显示,反馈和局部连接以紫色显示。所有这些处理最终汇聚于脊髓腹角运动神经元,即所谓的“最终共同通路”,它支配肌肉并引发运动。(这张图是从不同角度拍摄的大脑部分的合成视图。)
4.7 周围神经系统在解剖学上与中枢神经系统不同
4.8 记忆是一种复杂的行为,由不同于执行感觉或运动的结构介导
4.8.1 海马系统与最高层的多感觉皮层区域相互连接
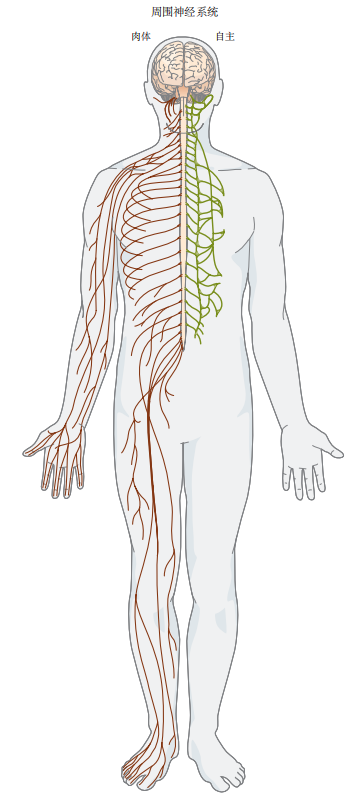
图 4.7.1:周围神经系统由躯体神经和自主神经分支组成。躯体神经分支负责将信息从皮肤传递到大脑,再从大脑传递到肌肉。自主神经系统调节非自主功能,包括心脏活动、肠道平滑肌和腺体。
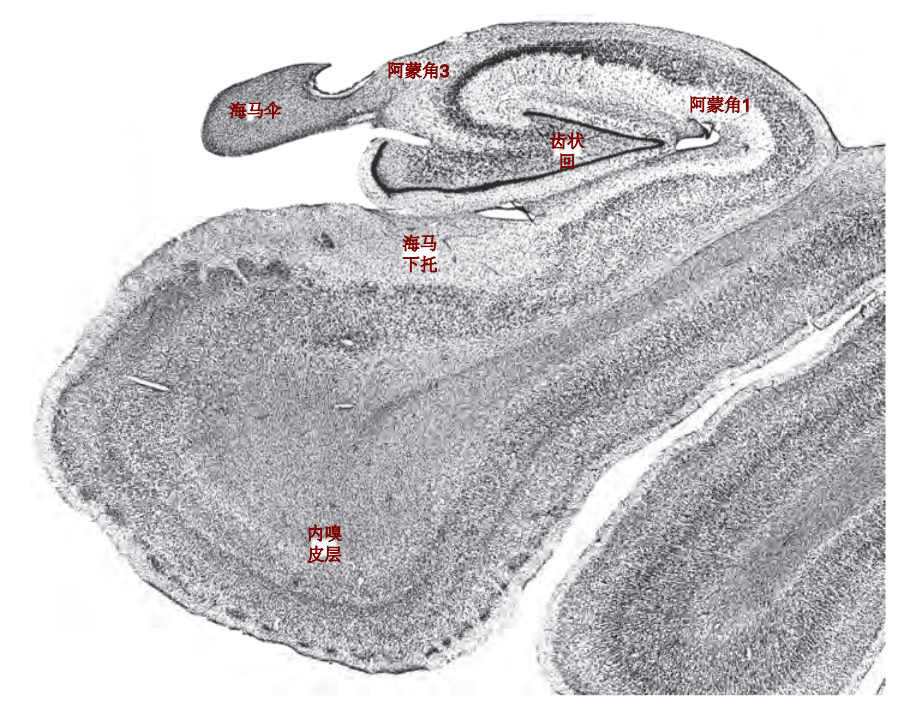
图 4.8.1:用尼氏染色法展示的人类海马体冠状切片,显示了海马结构的主要细胞体构造。
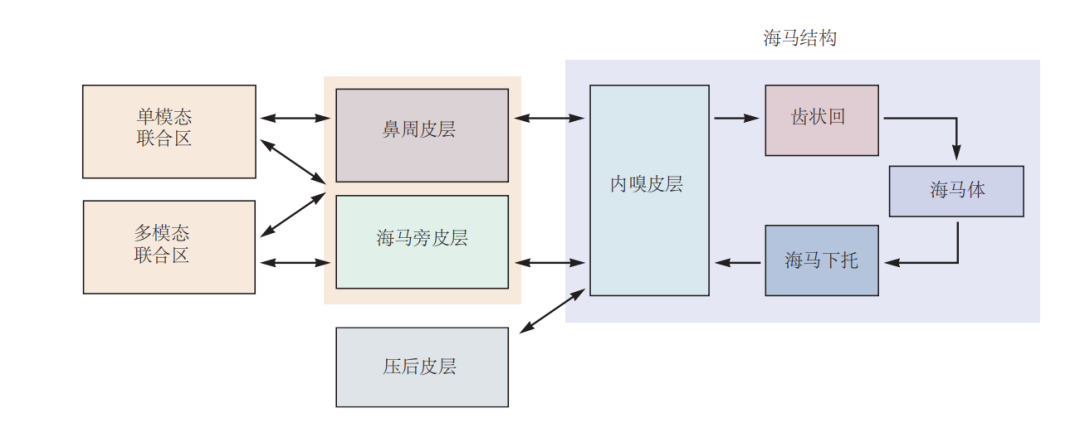
图 4.8.2:海马结构的层次连接组织。海马结构主要通过内嗅皮层接收来自多模态联合区(如鼻周皮层、海马旁皮层和后扣带皮层)的高度处理的感觉信息。
4.8.2 海马结构由几个不同但高度集成的回路组成
4.8.3 海马结构主要由单向连接组成
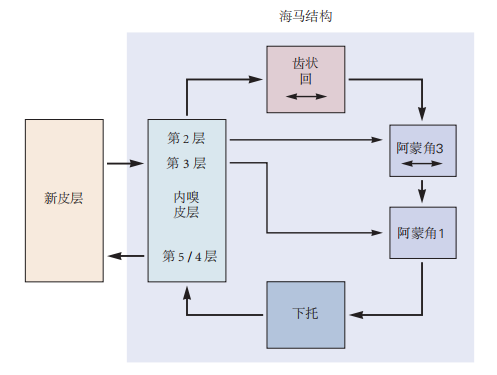
图 4.8.3:海马结构内部连接的简化图。该回路从内嗅皮层第2层的细胞开始,投射到齿状回,再投射到海马体的阿蒙角3区。接着,阿蒙角3投射到阿蒙角1,然后阿蒙角1投射到海马下托。当下托投射回内嗅皮层深层时,海马回路闭合。未显示的是从内嗅皮层到其接收多模态感觉信息的同一区域的反馈通路。
4.9 结语
本文翻译整理自Principles of Neural Science, Sixth Edition,翻译初稿来自于GitHub/OpenHUTB,本公众号进行进一步校对排版后发布,仅用于学习交流使用,如需购买原版书籍可点击阅读原文购买。
— THE END —
来源 | Principles of Neural Science, Sixth Edition
图文排版 | 刘玉娇
审核修改 | 王坤
(转载请保留此部分内容)

关注本号~

加入读者交流群~
(添加请备注单位姓名)

加入知识星球~
点亮赞与在看
让更多人看到
本篇文章来源于微信公众号:OncoLab