导 读
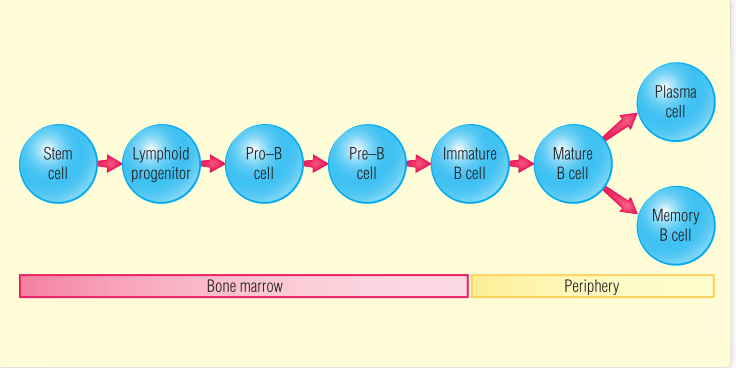
正如第6章和第11章所讨论的那样,B细胞表面的膜结合抗体分子构成负责识别抗原的B细胞受体(BCR)的关键部分。抗原特异性BCR经过一系列的发育过程才能最终表达,表达多种BCR的B淋巴细胞(尚未被选择)在骨髓中不断产生。在发育成熟之前,发育中的B细胞会经历一个被称为阴性选择的选择过程,以确保它们的抗原受体不识别自身抗原,否则自身反应性可能导致自身免疫疾病的发生(第28章)。在阴性选择中幸存下来的B细胞分散到外周淋巴器官(第13章),在那里它们可能会遇到具有特异性的外来抗原,然后被激活,最终分化为可以产生抗体的细胞。而没有遇到合适抗原的B细胞会在几周内死亡。本章和上图讲述了B细胞从造血干细胞到常规成熟B细胞的发育阶段。在这一章的最后,描述了两种非常规的B细胞群体,即非T细胞依赖性的B细胞和B1型B细胞。
B淋巴细胞的早期发育
干细胞转化为未成熟B细胞
B细胞来源于由造血干细胞分化形成的淋巴祖细胞(第12章)。人的一生都会产生B细胞,但B细胞的分化首先发生在胎儿肝脏,出生后不久便转移到骨髓(第12章和第13章)。在成人骨髓中,B细胞的发育成熟途径呈放射状,其中最不成熟的细胞靠近骨内膜表面,而较成熟的细胞集中在骨髓中央空间。未成熟的B细胞通过血窦流出骨髓,并迁移到外周(通常是脾脏或淋巴结)。它们在骨髓中的发育依赖于骨髓基质细胞提供的各种生长因子(第12章)。
从最早的组细胞–造血干细胞开始,B细胞的分化途径可细分为几个发育阶段。这些阶段是由免疫球蛋白(Ig)重链和轻链基因的重排以及细胞表面的分化特异性分子的表达来定义的,如Table14.1所示。
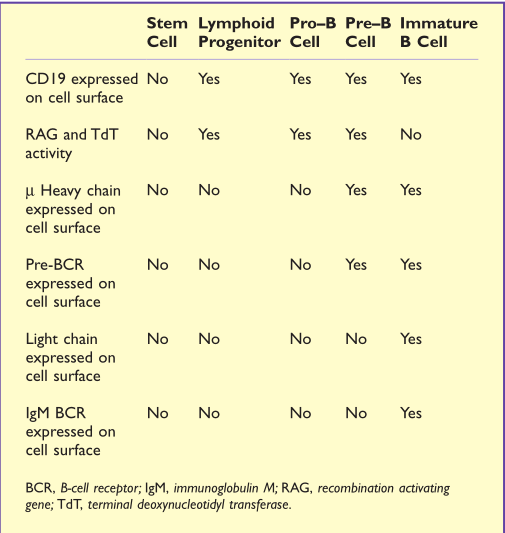
TABLE14.1分化特异性分子
最早确定的B系细胞被称为原B细胞(pro-B cell)。这种细胞是通过B系特有的表面标志物来识别的,例如CD19(共受体复合体的一部分)(第11章)。重组激活基因(RAG)在pro-B细胞中较活跃,Ig重链基因群中编码可变区的V基因片段(VH)、D基因片段(DH)和J基因片段(JH)进行重排,并产生重链。在这些细胞中, 末端脱氧核苷酸转移酶(TDT)也是活跃的,它可增加连接的多样性。
一旦表达了µ重链,该细胞就成为前B细胞(pre-B细胞)(见第6章)。µ重链(µH)与前B细胞受体(ψL)和µH链(第11章)一起在细胞表面表达,形成前B细胞受体(pre-BCR)的受体复合体。pre-BCR在pre-B细胞的增殖和促进其进一步发育的信号传导中起着重要作用。增殖促进了下一个主要阶段——晚期pre-B细胞的进一步发育。
在pre-B细胞阶段,V(D)J重组酶机制启动轻链基因重排。细胞首先重组κ轻链,如果重组不成功,它将产生λ轻链。当成对的轻链和重链多肽形成完整的IgM分子(BCR)在细胞表面表达时,表明细胞已发育成未成熟B细胞(Immature B),RAG和TDT酶则不会再进行转录(Fig14.1)。

Fig 14.1 在骨髓中产生表达BCR的未成熟B细胞
细胞经历上述成熟过程的同时也在分裂和增殖。最终可以产生大量的B细胞。另一方面,重组不能成功表达pre-BCR或BCR对细胞来说是致命的,这确保了只有BCR完好的B细胞才能在成熟过程中存活下来。
B细胞发育过程中的重组形成了受体多样性,但必然会产生一些具有自身反应性的抗原受体。这些受体很可能会识别一些自身抗原,表达这些受体的细胞需要被杀死或者抑制,这个过程被称为阴性选择。这一选择过程使得未成熟的B细胞在受到自身抗原的刺激时不会被激活,这种情况被称为耐受性。在本文中,自身抗原是指骨髓中存在的可以被BCR结合的大分子。这些分子可能是存在于健康细胞表面的分子,也可能是以溶液形式存在于细胞外基质中的分子。
细胞表面丰富的自身抗原可以广泛地与未成熟的BCR结合,诱导细胞凋亡,导致能够与自身抗原反应的克隆被清除。未成熟的B细胞对小且不能有效结合未成熟的BCR的可溶性蛋白的反应不同。当未成熟的B细胞暴露在高剂量的可溶性抗原下时,产生反应的B细胞会下调IgM的表达,随后对抗原刺激不产生应答,这种情况被称为失能。这些B细胞不会被杀死,某些情况下可以从失能中恢复过来。在阴性选择中幸存下来的B细胞会增加的IgM水平,并开始表达膜结合型IgD(Fig14.2)。
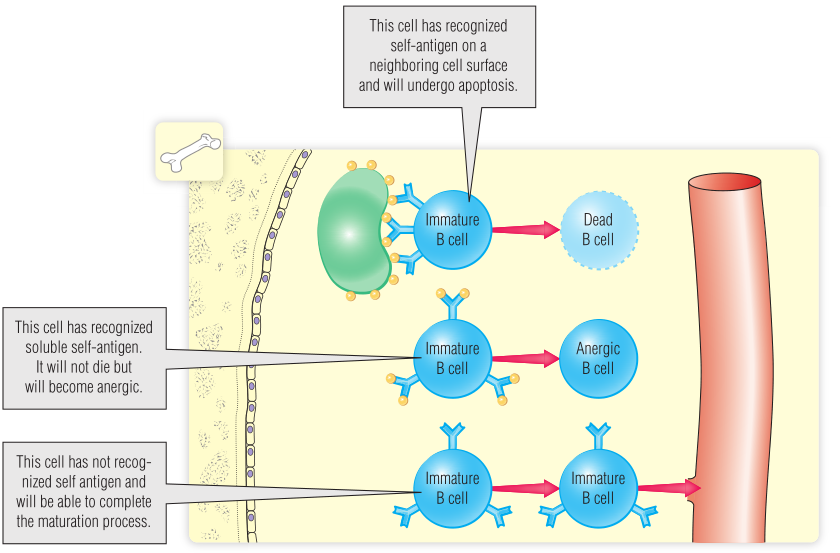
Fig 14.2 阴性选择确保只允许不识别自身抗原的未成熟B细胞离开骨髓
IgD在重链转录本选择性剪接后表达(第6章)。B细胞经过这一阶段后成为表达表面IgM和表面IgD的成熟B细胞。一旦B细胞在骨髓中完成早期成熟阶段,它就开始迁移到外周淋巴器官以完成其发育。已完全完成Ig基因重排但还没遇到非自身抗原的B细胞称为初始B细胞(naive B cells)。
成熟B细胞
额外的免疫耐受诱导
因为并不是所有的自身抗原都存在于骨髓中,进化机制可确保成熟的B细胞群对周围遇到的自身抗原具有耐受性。成熟的B细胞通常需要T细胞的帮助才能产生抗体。如果成熟的B细胞与抗原结合,而该抗原的特异性T细胞没有提供B细胞完全激活所需的信号(第11章和第16章),那么该细胞将被清除或失能。与未成熟的B细胞一样,受体结合取决于所遇到的抗原。多价抗原通常引起克隆清除,单价抗原则引起失能(Fig14.3)。然而,有一些B细胞已经进化成能够在没有T细胞帮助的情况下对多价多糖抗原产生特异性反应;这些不依赖胸腺(T细胞非依赖性)的B细胞将在本章后面进行讨论。
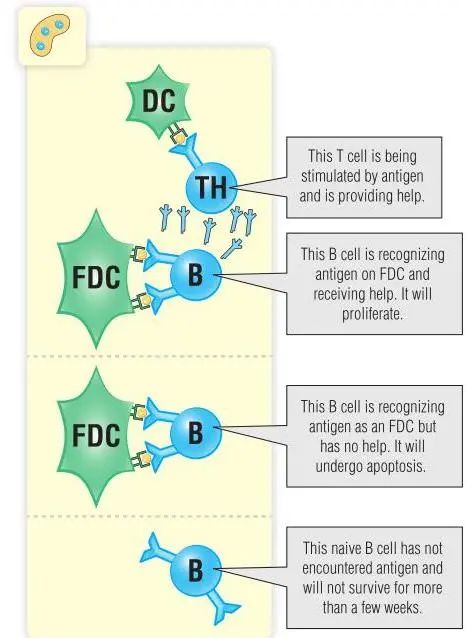
Fig 14.3 生发中心:初始B细胞识别抗原并接受T细胞的帮助
活化与抗体产生
对特定抗原做出反应的B细胞通常需要T细胞的帮助,并需要滤泡树突状细胞(FDCs)来提呈抗原。正如第13章所述,次级淋巴器官中的滤泡提供了一个环境,在那里抗原被集中并呈递给初始B细胞,并且初始 B细胞、FDCs和T细胞可以相互作用。每种类型的次级淋巴组织可捕获不同来源的抗原:如脾脏收集血液性抗原,淋巴结捕获进入淋巴系统中的抗原、粘膜相关淋巴组织(MALT),如肠道中的Peyer斑块和鼻咽中的扁桃体/腺样体,从周围的粘膜上皮细胞获得抗原。成熟的初始B细胞离开骨髓后会不断地通过血液循环到不同次级淋巴器官中的滤泡,直到遇到特定的抗原。缺乏目标抗原时,初始B细胞会穿过这一区域,并重新进入血液循环。
FDCs是一种特殊的抗原提呈细胞(APC),仅在淋巴滤泡中被发现。它们不表达Ⅱ类主要组织相容性复合体(MHC)分子,而是将抗原抗体复合物保留在其表面,附着在Fc受体或补体受体上。这些细胞具有在淋巴滤泡内形成的网络状长突起,与其他树突状细胞(DC)相似,但它们可能与骨髓中表达MHC II类分子的DC的来源不同。
一般来说,单纯的抗原不能激活初始B细胞。需要第二个辅助信号来实现B细胞的完全激活(第16章)。在抗原是蛋白质的情况下,第二个信号由已激活的T细胞提供,该T细胞带有抗原受体(TCR),可识别相邻APC显示的肽-MHC II复合物。只有通过这种形式的B细胞-T细胞相互协作才能启动免疫反应的抗原被称为胸腺依赖性抗原。
如果B细胞遇到它的目的抗原,BCR与抗原结合就会触发BCR抗原复合体内化进入B细胞。然后抗原被降解,抗原是蛋白质时,它被加工成肽-MHC II类复合物,随后呈递在细胞表面(第10章)。因此,B细胞可以向T细胞呈递抗原,T细胞则帮助邻近的B细胞进行激活作为回报。
生发中心的体细胞突变和类别转换
在抗原刺激和T细胞的帮助下,激活的B细胞大约以每6小时分裂一次形成生发中心,淋巴结内是由FDCs组成的浅染区域(见Fig13.3A和B)。在此期间,免疫球蛋白基因发生高频率的点突变而没有被纠正。这一独特的过程被称为体细胞突变,在任何其他类型的细胞中都不允许这种情况的发生。体细胞突变通常包括集中在重排可变区的外显子及其周围的单核苷酸替换。重链可变区外显子和轻链可变区外显子都可能是体细胞突变的靶点。引入的突变可能会影响BCR与抗原的结合情况。那些BCR与抗原高亲和力的B细胞(Fig14.4)从FDCs和生发中心T细胞获得存活信号(阳性选择),而那些不能结合抗原的B细胞很快凋亡。一旦感染得到控制,抗原水平开始下降。阳性选择的B细胞可能会经历额外的几轮增殖、体细胞突变和抗原选择。其结果是产生高亲和力抗体的B细胞存活。由此形成的亲和力成熟是抗体质量随时间提高的原因之一。例如,Fig2.1显示了接种疫苗后抗体水平是如何提高的。这在一定程度上是因为更多的B细胞产生抗体,但也很大程度上是因为抗体产生的亲和力随着时间的推移而增加。免疫球蛋白的数量和抗体结合的质量(亲和力)都会影响抗体水平。
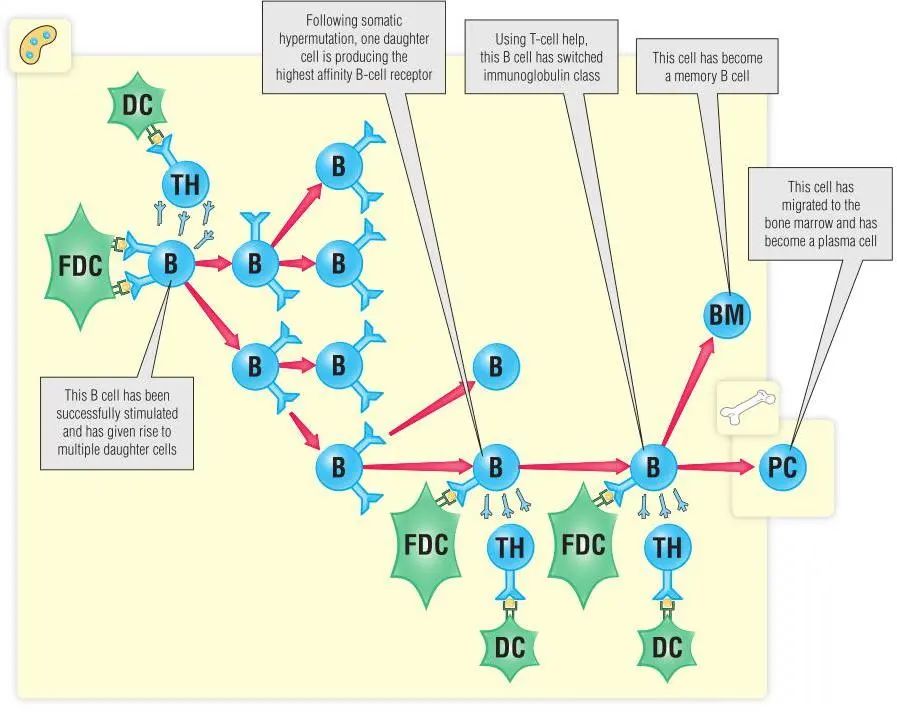
Fig 14.4 发育B细胞可能有几种不同的命运
生发中心发生的第二个重要发育过程是Ig的类别转换。在成熟过程中,B细胞表达IgM和IgD,但只分泌IgM。许多情况下产生其他类别的抗体是很重要的。例如,IgA在应对粘膜感染时是有帮助的。而这需要T细胞为Ig类别转换提供特定类型的帮助(第6章和第16章)
成熟过程的任何阶段B细胞都可能离开生发中心,最终分化为浆细胞或成为再循环的记忆B细胞,这两个步骤都需要B细胞的帮助。骨髓中的浆细胞可能会分泌大量抗体并且至少持续数周。
记忆B细胞不分泌抗体,但可以在之后遇到抗原攻击时迅速重新激活。再看看Fig2.4,在第二次,特别是第三次加强接种乙肝疫苗后,抗体水平迅速上升。这是典型的记忆B细胞反应。
胸腺非依赖性抗原
如前所述,对多肽抗原有反应的B细胞需要两种信号才能完全激活:与BCR的结合和T细胞的帮助。然而,有些抗原能够在没有T细胞帮助的情况下直接激活B细胞,因此它们被称为胸腺非依赖性(TI)抗原,脾脏中有大量的TI B细胞。B细胞直到儿童后期才完全发育,这是儿童容易反复受到细菌感染的原因之一。TI抗原包括重复聚合物(如细菌多糖)和某些细菌细胞壁成分(如脂多糖)。这些抗原都有多个重复基序,并与几个BCR结合,这样就克服了对第二个信号的需要(Fig14.5)。

Fig 14.5 T非依赖性(TI)抗原通过结合多个B细胞受体(BCR)来克服对T细胞帮助的需求
TI抗原通常是细菌起源的,TI反应提供了一种对细菌病原体产生早期和特异性抗体反应的方法,它能抵御细菌病原体的侵袭,这些病原体可以迅速繁殖,并使免疫系统不堪重负。然而,由于T细胞通常不被动员,在TI反应期间产生的抗体谱系是有限的,因为不会诱导促进亲和力成熟的T细胞依赖事件、Ig的类别转换和记忆B细胞的产生。
TI B细胞有两个重要的意义。首先,开发出能够诱导对有荚膜的细菌产生保护作用的疫苗,如嗜血杆菌和肺炎球菌(肺炎链球菌)。这些细菌会导致大量儿童死亡。然而,仅仅基于TI抗原多糖的疫苗不能刺激T细胞的帮助,因为它们不包含可以分解成多肽的蛋白质。因此在当用作疫苗时,多糖只能诱导免疫记忆力差的低亲和力IgM抗体。蛋白质结合多糖疫苗已经成功地克服了这个问题(第16章)。
接受过脾切除术的人受侵袭性感染的风险很高。正如第13章所描述的,部分原因是这些患者失去了大量能够过滤血液的脾脏巨噬细胞。感染风险高的第二个原因是TI B细胞的丧失,而这可以通过终生使用抗生素和结合多糖疫苗来进行治疗。
B1 细胞与天然抗体
B1细胞由未成熟的B细胞前体细胞发育而来,但它们在几个重要方面与传统的B细胞不同。首先,B1 细胞位于不寻常的部位,如腹膜,而不是传统的淋巴器官。第二,B1 细胞表达不同的细胞表面标志。例如,许多(但不是全部)B1细胞表达CD5的表面标记,这在传统的B细胞上是不存在的。
与从骨髓前体细胞不断更新的传统B细胞不同,成人的B1细胞通过外周组织中表面携带IgM(SIgM+)的细胞的持续分裂来进行补充,从而确保在其产生停止后的很长一段时间B1群体数量保持不变。这一特性可能会促使B1细胞(特别是CD5+细胞)成为一种相对常见的B细胞肿瘤的常见来源,称为慢性淋巴细胞白血病(第35章)。
由于少数VH基因的优先使用,B1细胞受体谱系受到很大限制,并且由于这些细胞中缺乏TDT的表达,插入多样性也是有限的。B1细胞上表达的BCR通常对细菌抗原(如多糖)具有反应性,并且它们经常表现出多重特异性,即与多种抗原发生交叉反应的能力。因此,B1细胞在生命早期对抗病原体功能提供了重要的支持,直到成年细胞发育。在这种情况下,B1细胞类似于TI B细胞(Table14.2)。
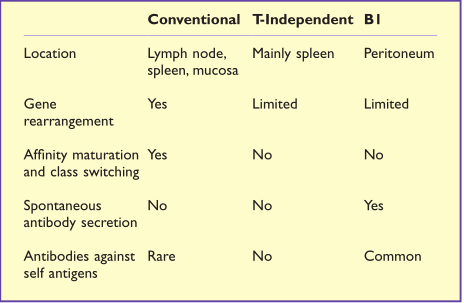
Table 14.2 B细胞的分类
然而,B1细胞在不受抗原刺激的情况下产生抗体时,抗体经常与自身抗原发生交叉反应。自发产生的自身反应性抗体被称为天然抗体,有时可能会引起自身免疫,如第28章。
最后,由于B1 细胞只使用有限的基因进行重排,不会随着时间的推移(通过亲和力成熟或类别转换)来完善它们的反应,也不会形成免疫记忆,所以它们可能与先天性免疫系统有更多的共同点。
BOX 14.1急性淋巴细胞性白血病
一名7岁的女孩,能量摄入不足和体重减轻3周。脸色苍白,淋巴结肿大,腿部有瘀点。血常规结果示血红蛋白和血小板计数较低(TABLE14.3)。白细胞数增多,但几乎所有的白细胞都是母细胞(非常不成熟的白细胞)。血液学专家观察这些细胞后认为它们可能是未成熟的淋巴细胞,并采用流式细胞术来确定(Table14.4)。由于CD19呈阳性,流式细胞术显示异常细胞是B细胞,并且这些B细胞表达TDT,表明它们非常不成熟。这对急性淋巴细胞白血病(ALL)的诊断非常有意义。这一诊断是通过对骨髓样本的检查来确认的。
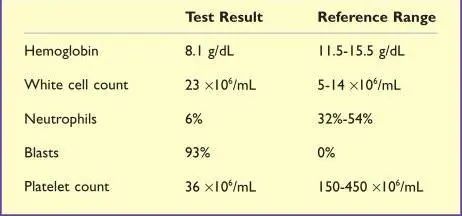
Table 14.3 全血细胞计数
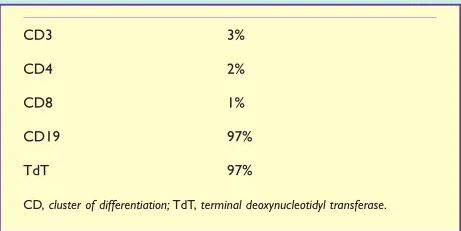
Table 14.4 流式细胞术
ALL是大多数发达国家最常见的儿童癌症,80%患有ALL的儿童可以治愈。恶性细胞起源于原始B细胞(见Fig14.1)。虽然暴露于辐射是ALL发病的已知危险因素之一,但大多数ALL儿童似乎并不是因暴露于辐射下患病的。
本文由Oncolab实验室微信公众号翻译整理自Immunology for Medical Students,仅用于学习交流使用,如需购买原版书籍可点击阅读原文购买。
— THE END —
▉ 往期精彩内容推荐

长按上方二维码即可关注本号~
本篇文章来源于微信公众号:OncoLab