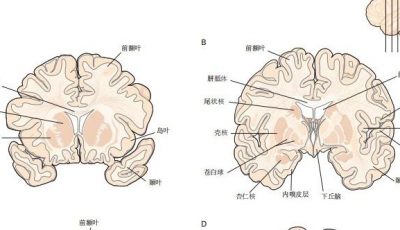
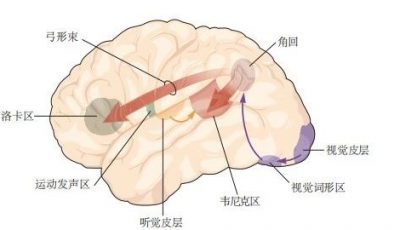
5.2 神经回路基序为信息处理提供了基本逻辑
5.2.2 小脑中不同的神经元表征为学习提供了基础
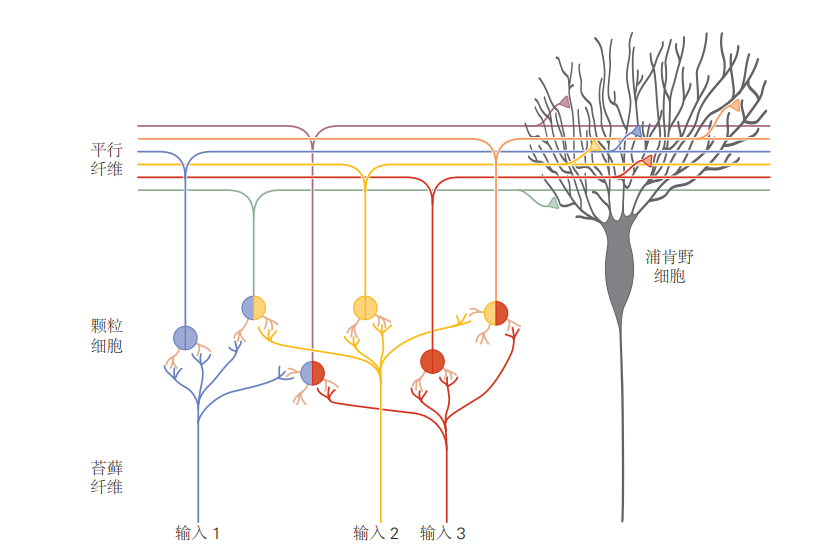
图 5.2.3:小脑接收来自大脑和脊髓多个区域的输入。这些输入统称为苔藓纤维,在大量颗粒细胞中进行重新编码。这个过程是发散连接的一个例子,它允许输入信号的多种可能组合。浦肯野细胞的树突接收来自数十万个颗粒细胞的会聚输入,这些细胞通过其轴突(称为平行纤维)中继信号。浦肯野细胞突触的平行纤维具有可塑性,这被认为是运动学习以及可能的其他学习形式的重要机制。
5.2.3 循环回路是持续活动和整合的基础
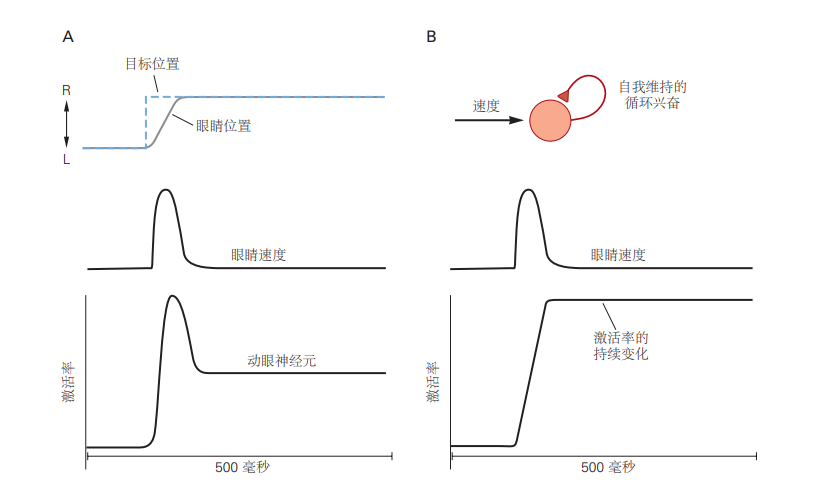
图 5.2.4:维持眼睛位置需要循环回路和持续的神经活动。A. 上图:扫视眼球运动包括眼球速度的快速变化,将目标带回凝视中心。随后,眼睛位置持续调整,以将中央凹对准目标。蓝色虚线表示目标位置,灰色线表示眼球运动及随后的注视。下图:动眼神经元表现出与眼球速度相关的短暂活动以及与眼睛位置相关的持续活动。B. 反复的兴奋可以解释如何通过类似于数学积分的过程,将短暂的脉冲输入(如眼速信号)转化为持续的放电率变化。
5.3 学习和记忆取决于突触可塑性
5.3.1 突触输入的主导模式可以通过赫布可塑性来识别
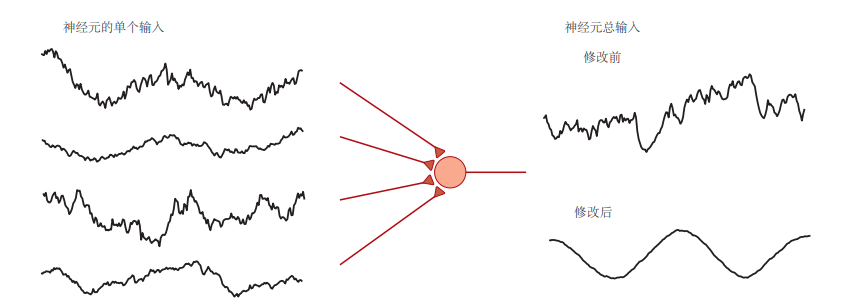
图 5.3.1:赫布可塑性可以识别神经元的相关输入信号。在这个例子中,一个神经元接收100个输入,其中显示了4个输入的激活率(左图)。每个输入速率都包含一定的噪声,但其中还包含正弦信号。输入速率通过突触强度(棕色三角形)加权后求和,生成神经元的总输入(右图)。在赫布可塑性作用之前,突触权重是随机的,导致总输入表现出噪声轨迹;经过赫布可塑性调整后,总输入中揭示了潜在的正弦信号。
5.3.2 小脑的突触可塑性在运动学习中起着关键作用
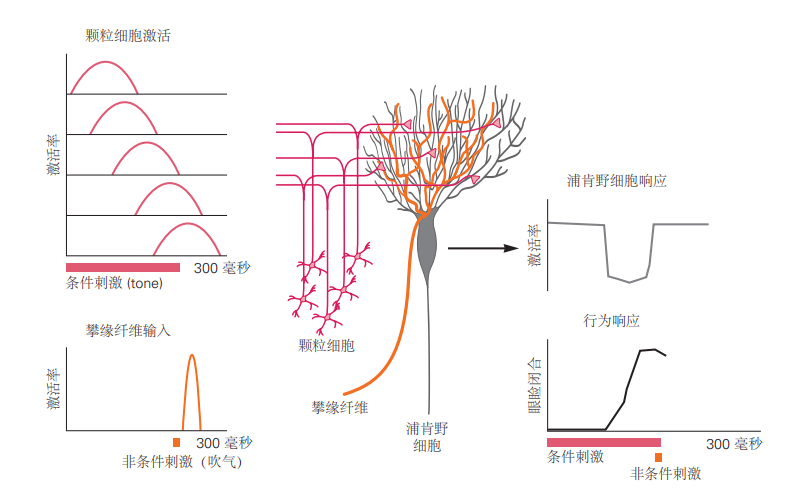
图 5.3.2:小脑在眨眼条件反射中的假设作用。有关条件刺激和非条件刺激的信息分别通过苔藓纤维和攀爬纤维通路传递。在非条件刺激呈现之前活跃的颗粒细胞突触逐渐因攀爬纤维输入引发的长期抑制而减弱。这有助于浦肯野细胞的暂停激活,恰好在非条件刺激发生前出现。由于浦肯野细胞是抑制性的,这种暂停会激活小脑核和红核中的下游神经元,从而驱动眼睑闭合。
5.4 本章要点回顾
本文翻译整理自Principles of Neural Science, Sixth Edition,翻译初稿来自于GitHub/OpenHUTB,本公众号进行进一步校对排版后发布,仅用于学习交流使用,如需购买原版书籍可点击阅读原文购买。
— THE END —
来源 | Principles of Neural Science, Sixth Edition
图文排版 | 曾世纪
审核修改 | 王坤
(转载请保留此部分内容)

关注本号~

加入读者交流群~
(添加请备注单位姓名)

加入知识星球~
点亮赞与在看
让更多人看到
本篇文章来源于微信公众号:OncoLab